

Toward resolution of ambiguity for the unfolded state
- PMID: 18469075
- PMCID: PMC2440439
- DOI: 10.1529/biophysj.107.121855
Toward resolution of ambiguity for the unfolded state
Abstract
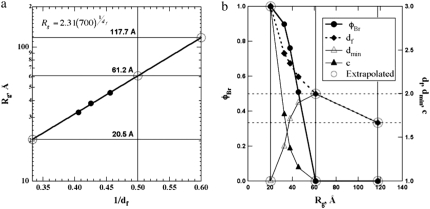
The unfolded states in proteins and nucleic acids remain weakly understood despite their importance in folding processes; misfolding diseases (Parkinson's and Alzheimer's); natively unfolded proteins (as many as 30% of eukaryotic proteins, according to Fink); and the study of ribozymes. Research has been hindered by the inability to quantify the residual (native) structure present in an unfolded protein or nucleic acid. Here, a scaling model is proposed to quantify the molar degree of folding and the unfolded state. The model takes a global view of protein structure and can be applied to a number of analytic methods and to simulations. Three examples are given of application to small-angle scattering from pressure-induced unfolding of SNase, from acid-unfolded cytochrome c, and from folding of Azoarcus ribozyme. These examples quantitatively show three characteristic unfolded states for proteins, the statistical nature of a protein folding pathway, and the relationship between extent of folding and chain size during folding for charge-driven folding in RNA.
Figures



References
-
- Chaudhuri, T. K., and S. Paul. 2006. Protein-misfolding diseases and chaperone-based therapeutic approaches. FEBS J. 273:1331–1349. - PubMed
-
- Nissen, P., J. Hansen, N. Ban, P. B. Moore, and T. A. Steitz. 2000. The structural basis of ribosome activity in peptide bond synthesis. Science. 289:920–930. - PubMed
-
- Tanford, C. 1968. Protein denaturation. Adv. Protein Chem. 23:121–282. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
