

Utilization of the least shrew as a rapid and selective screening model for the antiemetic potential and brain penetration of substance P and NK1 receptor antagonists
- PMID: 18471804
- PMCID: PMC2486262
- DOI: 10.1016/j.brainres.2008.03.077
Utilization of the least shrew as a rapid and selective screening model for the antiemetic potential and brain penetration of substance P and NK1 receptor antagonists
Abstract
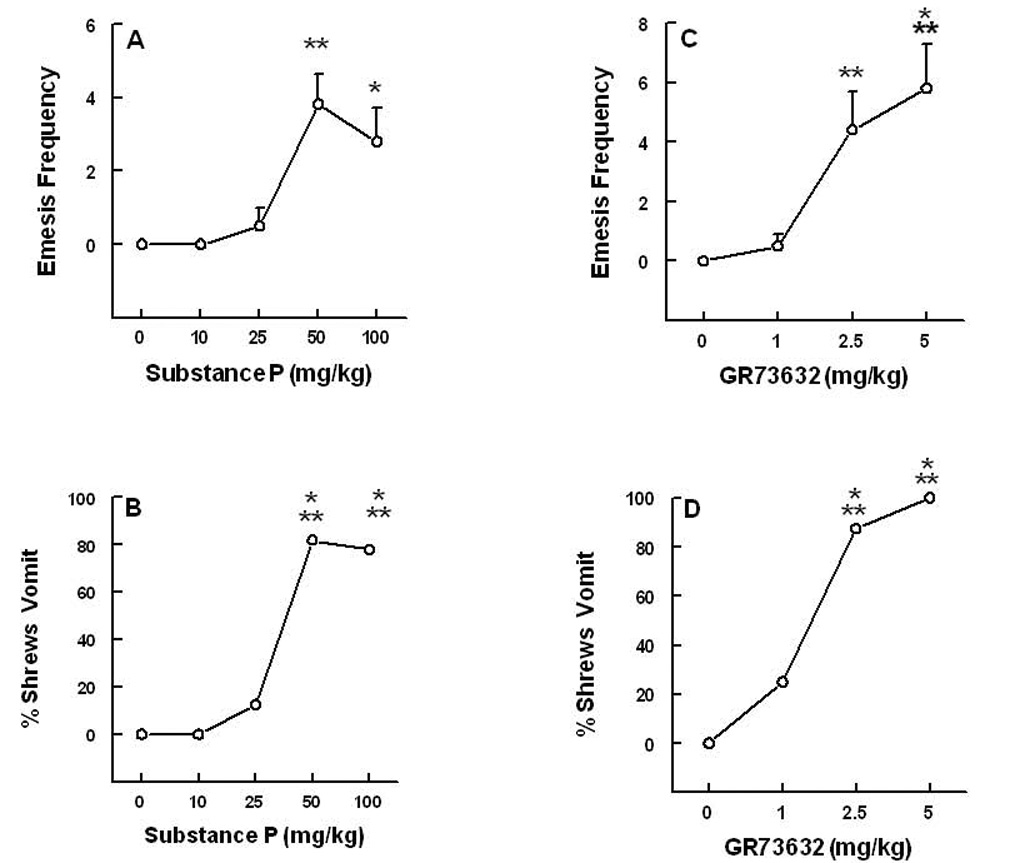
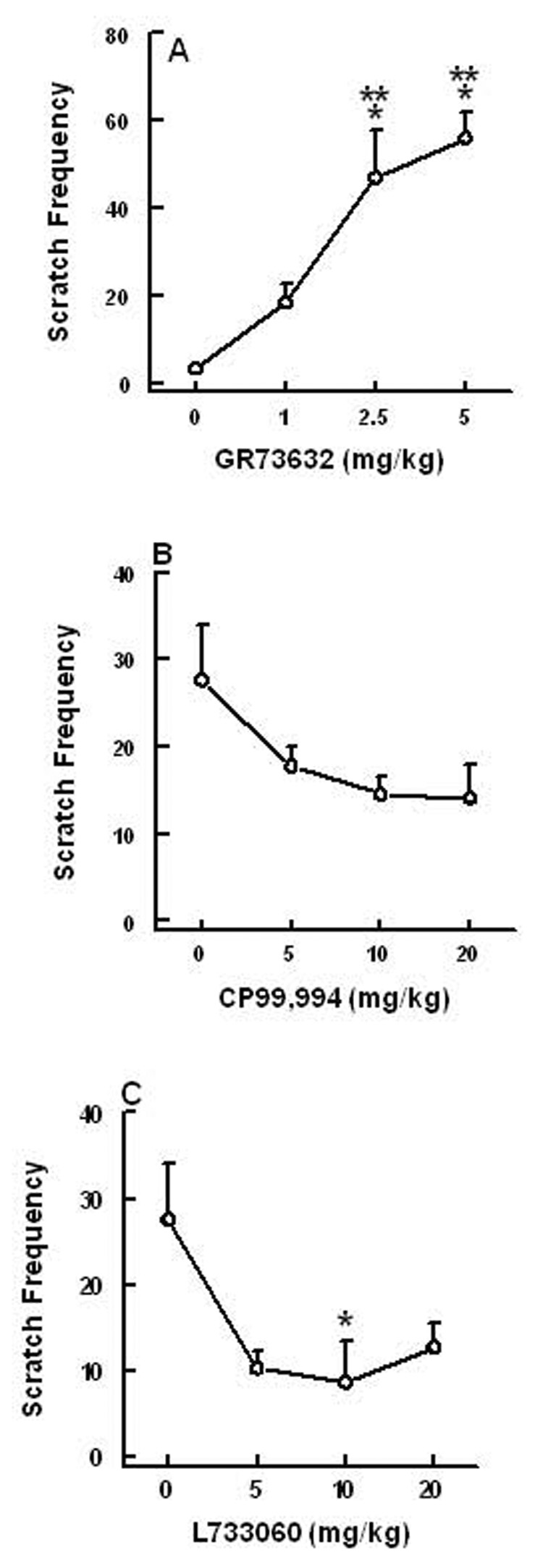
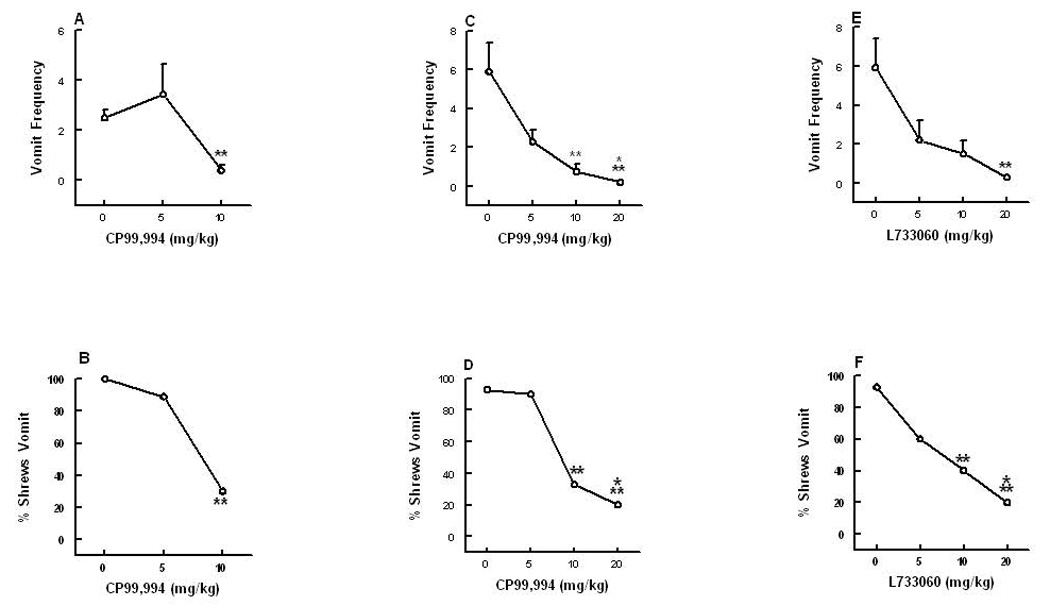
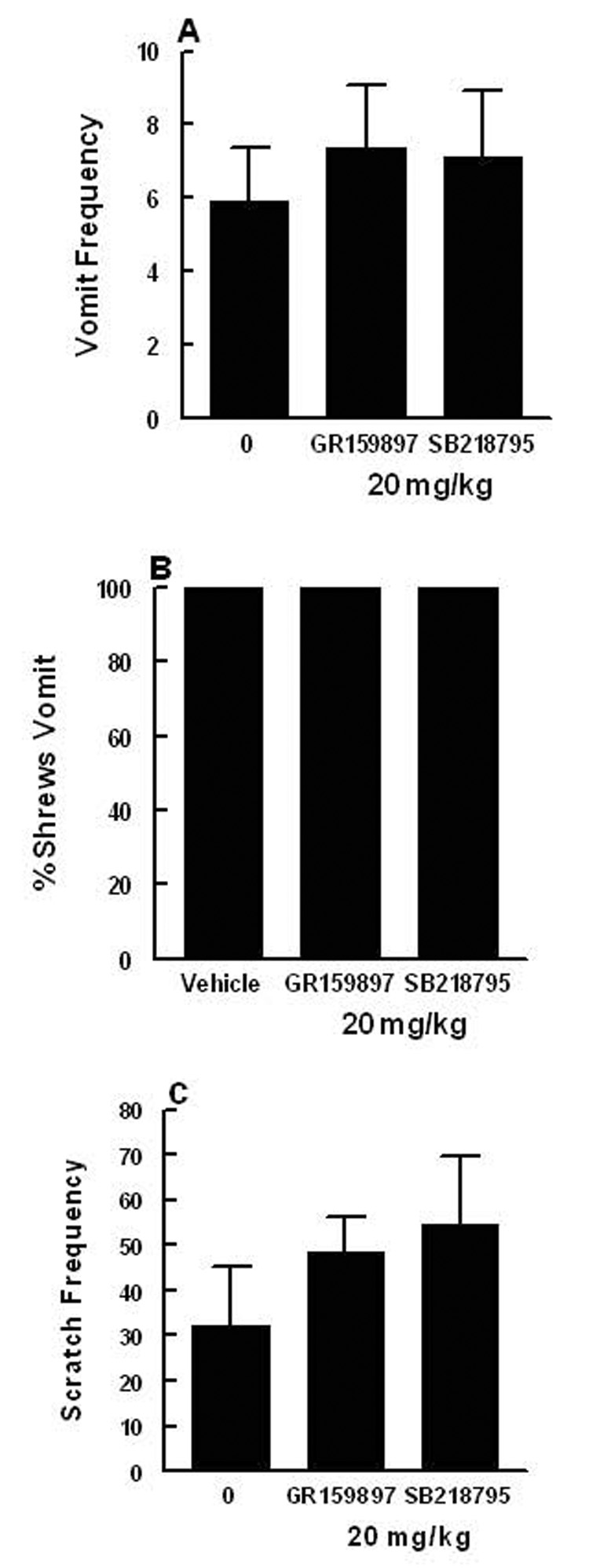
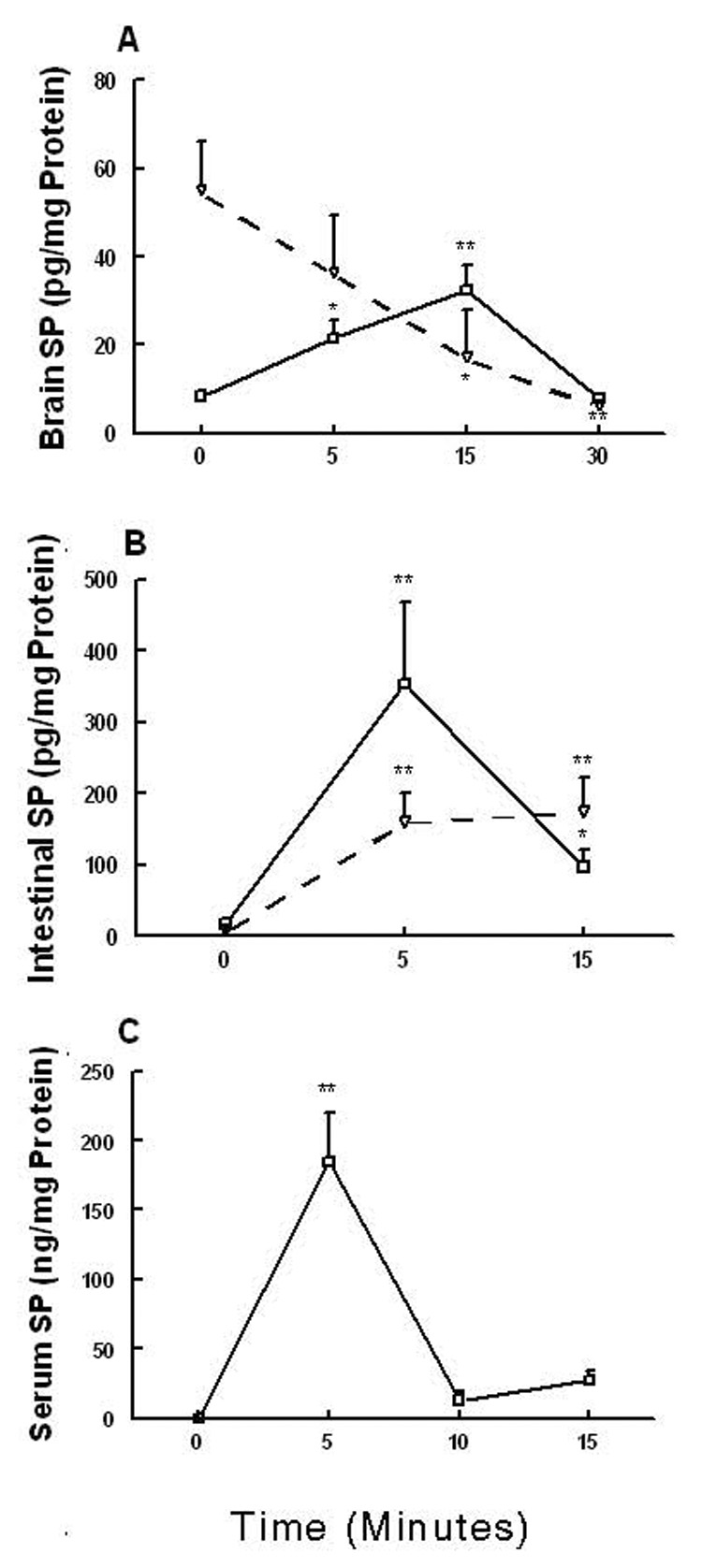
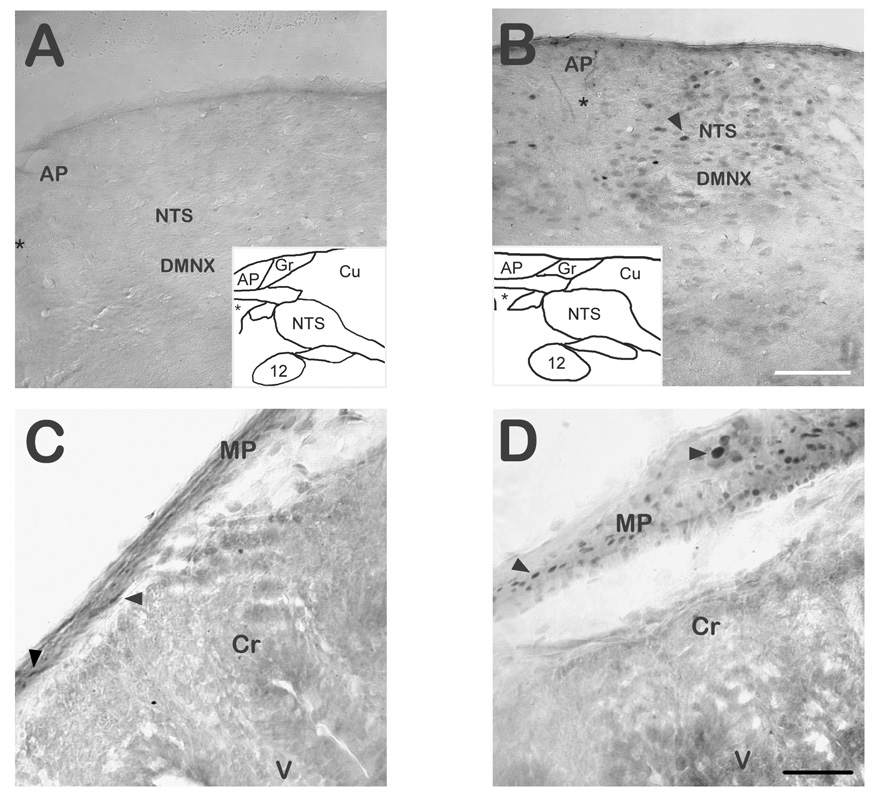
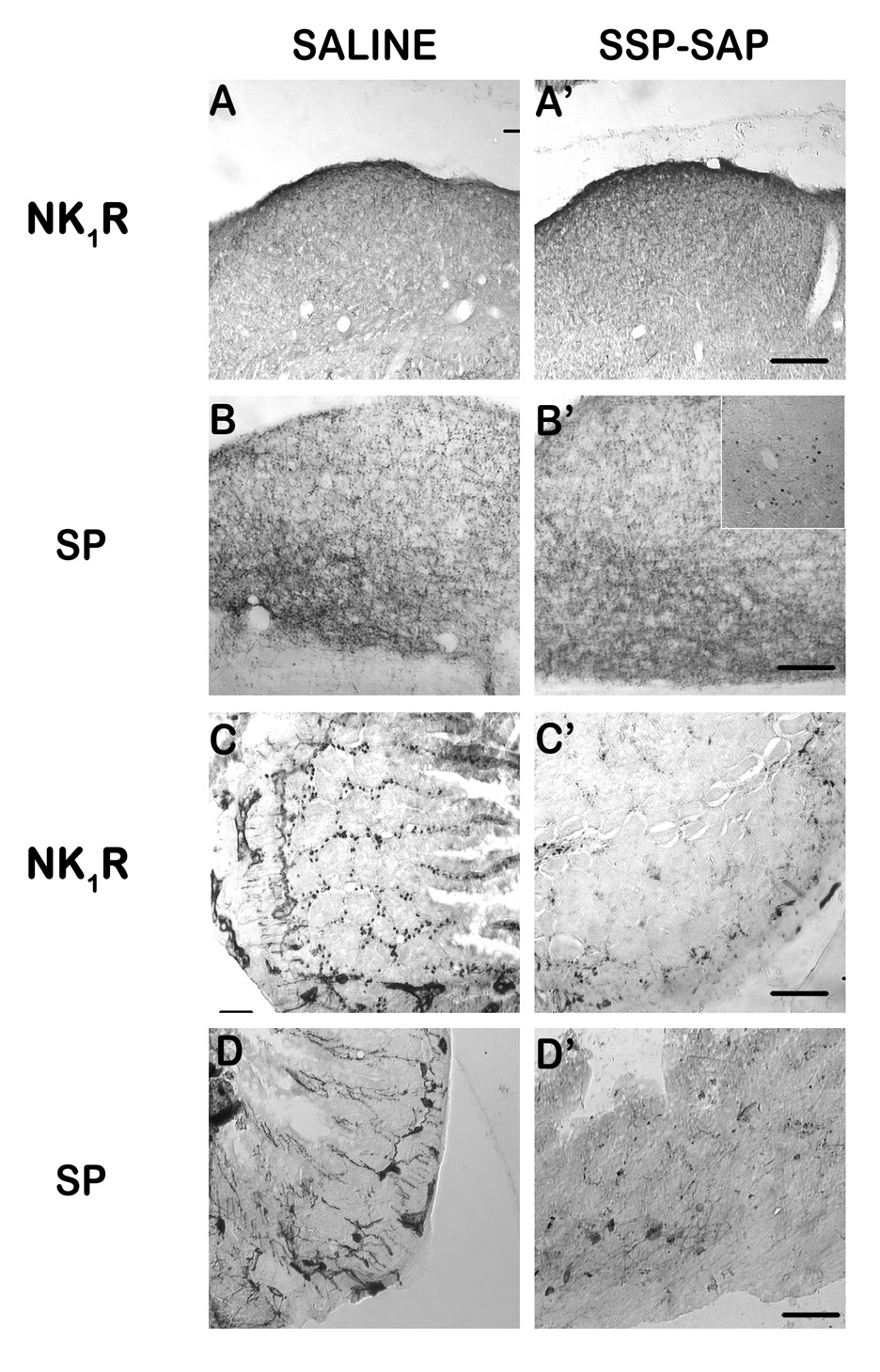
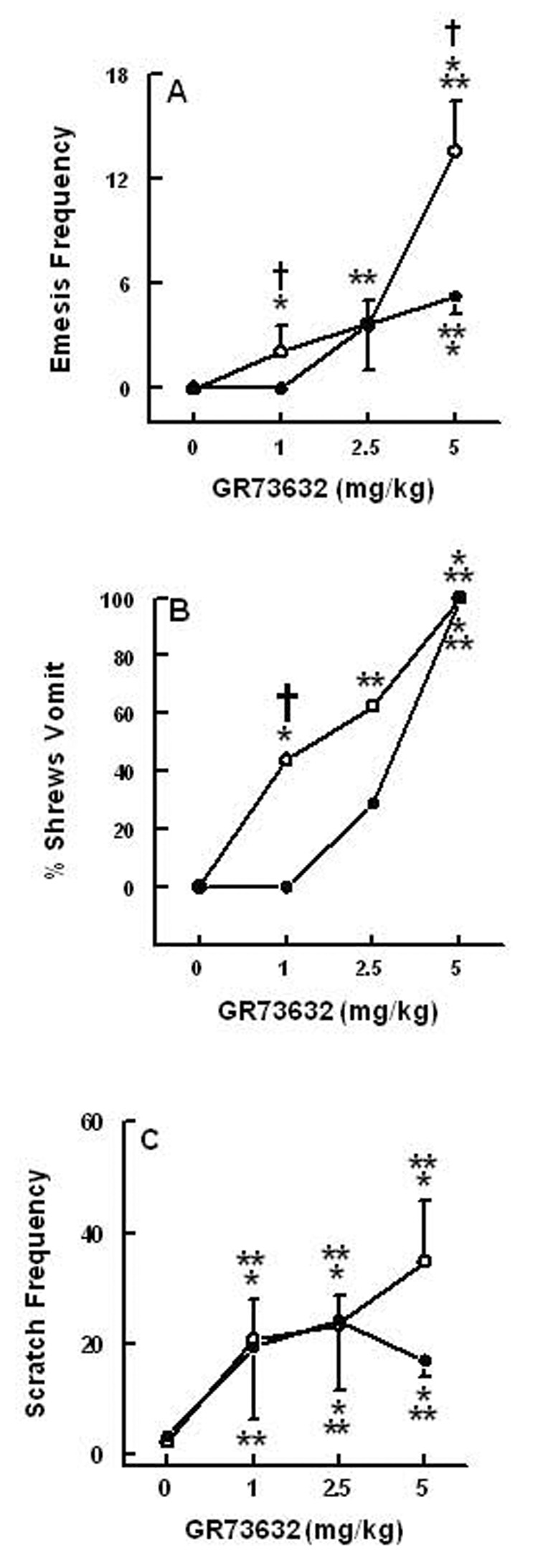
Substance P (SP) is thought to play a cardinal role in emesis via the activation of central tachykinin NK1 receptors during the delayed phase of vomiting produced by chemotherapeutics. Although the existing supportive evidence is significant, due to lack of an appropriate animal model, the evidence is indirect. As yet, no study has confirmed that emesis produced by SP or a selective NK1 receptor agonist is sensitive to brain penetrating antagonists of either NK1, NK2, or NK3 receptors. The goals of this investigation were to demonstrate: 1) whether intraperitoneal (i.p.) administration of either SP, a brain penetrating (GR73632) or non-penetrating (e.g. SarMet-SP) NK1 receptor agonist, an NK2 receptor agonist (GR64349), or an NK3 receptor agonist (Pro7-NKB), would induce vomiting and/or scratching in the least shrew (Cryptotis parva) in a dose-dependent manner; and whether these effects are sensitive to the above selective receptor antagonists; 2) whether an exogenous emetic dose of SP (50 mg/kg, i.p.) can penetrate into the shrew brain stem and frontal cortex; 3) whether GR73632 (2.5 mg/kg, i.p.)-induced activation of NK1 receptors increases Fos-measured neuronal activity in the neurons of both brain stem emetic nuclei and the enteric nervous system of the gut; and 4) whether selective ablation of peripheral NK1 receptors can affect emesis produced by GR73632. The results clearly demonstrated that while SP produced vomiting only, GR73632 caused both emesis and scratching behavior dose-dependently in shrews, and these effects were sensitive to NK1-, but not NK2- or NK3-receptor antagonists. Neither the selective, non-penetrating NK1 receptor agonists, nor the selective NK2- or NK3-receptor agonists, caused a significant dose-dependent behavioral effect. An emetic dose of SP selectively and rapidly penetrated the brain stem but not the frontal cortex. Systemic GR73632 increased Fos expression in the enteric nerve plexi, the medial subnucleus of nucleus tractus solitarius, and the dorsal motor nucleus of the vagus, but not the area postrema. Ablation of peripheral NK1 receptors attenuated the ability of GR73632 to induce a maximal frequency of emesis and shifted its percent animals vomiting dose-response curve to the right. The NK1-ablated shrews exhibited scratching behavior after systemic GR73632-injection. These results, for the first time, affirm a cardinal role for central NK1 receptors in SP-induced vomiting, and a facilitatory role for gastrointestinal NK1 receptors. In addition, these data support the validation of the least shrew as a specific and rapid behavioral animal model to screen concomitantly both the CNS penetration and the antiemetic potential of tachykinin NK1 receptor antagonists.
Figures
References
-
- Andrews PLR, Rudd JA. Handbook of Experimental Pharmacology. Vol. 164. Berlin: Springer-Verlag; 2004. The role of tachykinins and the tachykinin NK1 receptor in nausea and emesis; pp. 359–440.
-
- Bailey CP, Maubach KA, Jones RS. Neurokinin-1 receptors in the rat nucleus tractus solitarius: pre- and postsynaptic modulation of glutamate and GABA release. Neuroscience. 2004;127:467–479. - PubMed
-
- Begley DJ. The blood-brain barrier: principles for targeting peptides and drugs to the central nervous system. J Pharm Pharmacol. 1996;48:136–146. - PubMed
-
- Boissonade FM, Davison JS. Effect of vagal and splanchnic nerve section on Fos expression in ferret brain stem after emetic stimuli. Am J Physiol. 1996;271:R228–R236. - PubMed
-
- Boissonade FM, Davison JS, Egizii R, Lucier GE, Sharkey KA. The dorsal vagal complex of the ferret: anatomical and immunohistochemical studies. Neurogastroenterol Motil. 1996;8:255–272. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
