

Twist1 homodimers enhance FGF responsiveness of the cranial sutures and promote suture closure
- PMID: 18471809
- PMCID: PMC2605972
- DOI: 10.1016/j.ydbio.2008.03.037
Twist1 homodimers enhance FGF responsiveness of the cranial sutures and promote suture closure
Erratum in
- Dev Biol. 2012 Jun 15;366(2):434
Abstract
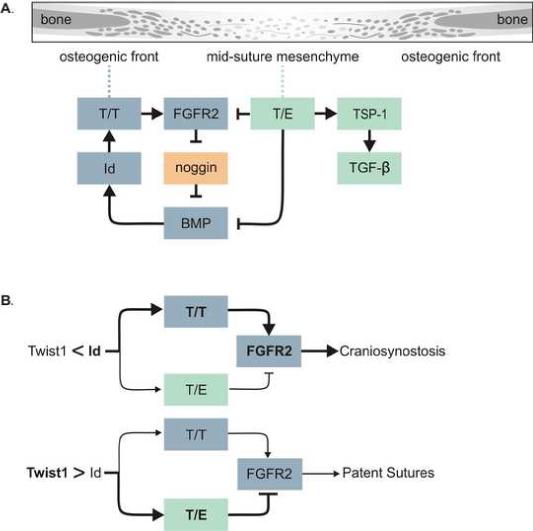
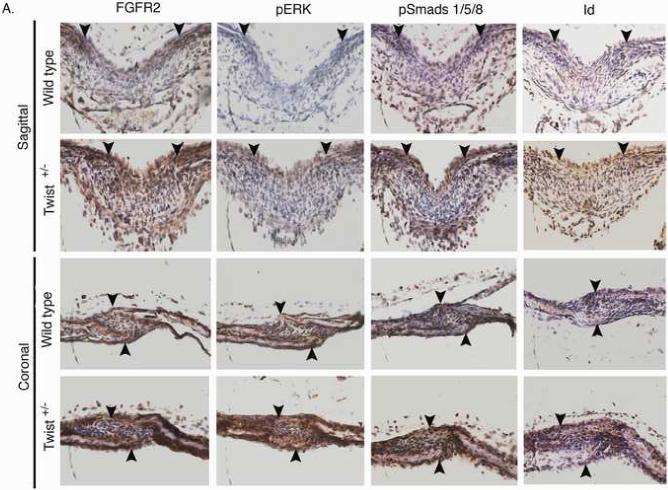
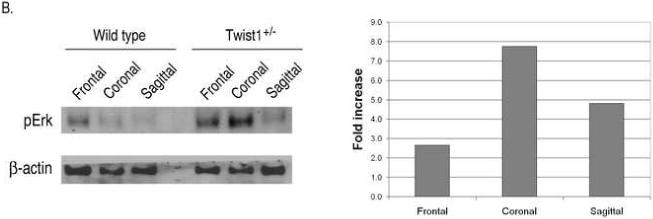
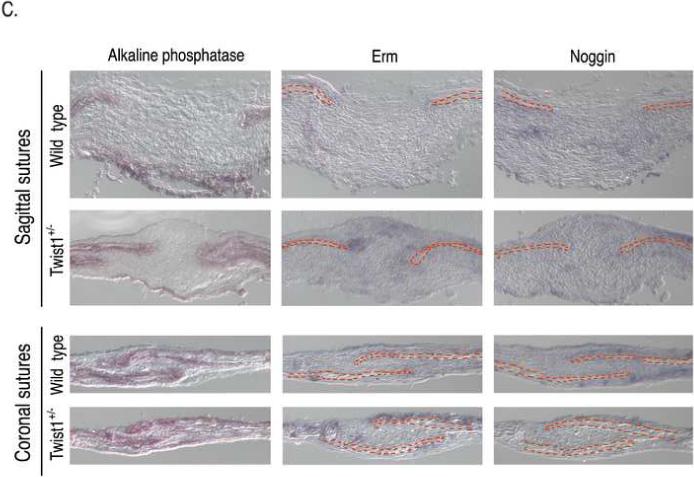
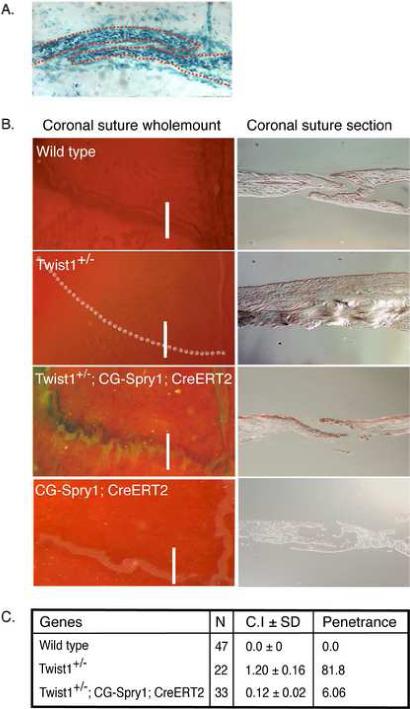
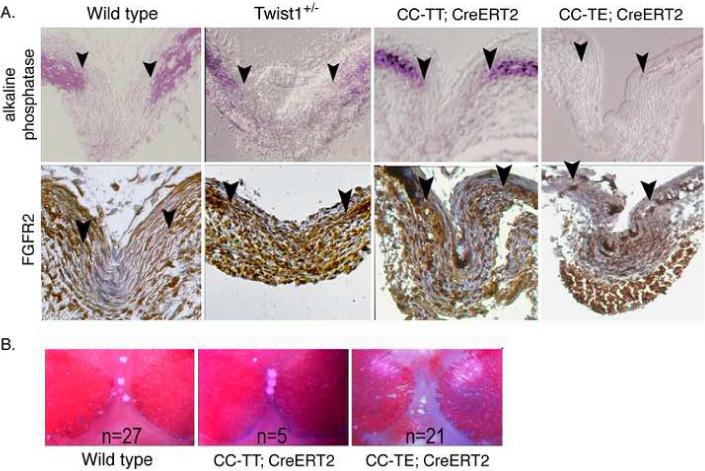
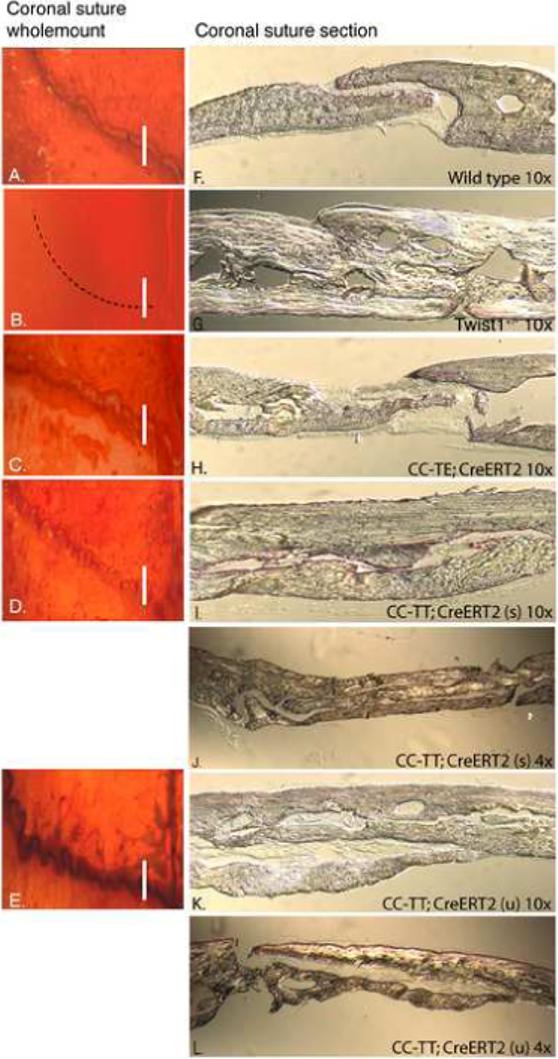
Haploinsufficiency of the transcription factor TWIST1 is associated with Saethre-Chotzen Syndrome and is manifested by craniosynostosis, which is the premature closure of the calvaria sutures. Previously, we found that Twist1 forms functional homodimers and heterodimers that have opposing activities. Our data supported a model that within the calvaria sutures Twist1 homodimers (T/T) reside in the osteogenic fronts while Twist1/E protein heterodimers (T/E) are in the mid-sutures. Twist1 haploinsufficiency alters the balance between these dimers, favoring an increase in homodimer formation throughout the sutures. The data we present here further supports this model and extends it to integrate the Twist1 dimers with the pathways that are known to regulate cranial suture patency. This data provides the first evidence of a functional link between Twist1 and the FGF pathway, and indicates that differential regulation of FGF signaling by T/T and T/E dimers plays a central role in governing cranial suture patency. Furthermore, we show that inhibition of FGF signaling prevents craniosynostosis in Twist1(+/-) mice, demonstrating that inhibition of a signaling pathway that is not part of the initiating mutation can prevent suture fusion in a relevant genetic model of craniosynostosis.
Figures

References
-
- Annes JP, et al. Making sense of latent TGFbeta activation. J Cell Sci. 2003;116:217–24. - PubMed
-
- Bialek P, et al. A twist code determines the onset of osteoblast differentiation. Dev Cell. 2004;6:423–35. - PubMed
-
- Brent AE, Tabin CJ. FGF acts directly on the somitic tendon progenitors through the Ets transcription factors Pea3 and Erm to regulate scleraxis expression. Development. 2004;131:3885–96. - PubMed
-
- Carver EA, et al. Craniosynostosis in Twist heterozygous mice: A model for Saethre-Chotzen syndrome. Anat Rec. 2002;268:90–2. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
