

Myosin filament 3D structure in mammalian cardiac muscle
- PMID: 18472277
- PMCID: PMC2531245
- DOI: 10.1016/j.jsb.2008.03.011
Myosin filament 3D structure in mammalian cardiac muscle
Abstract
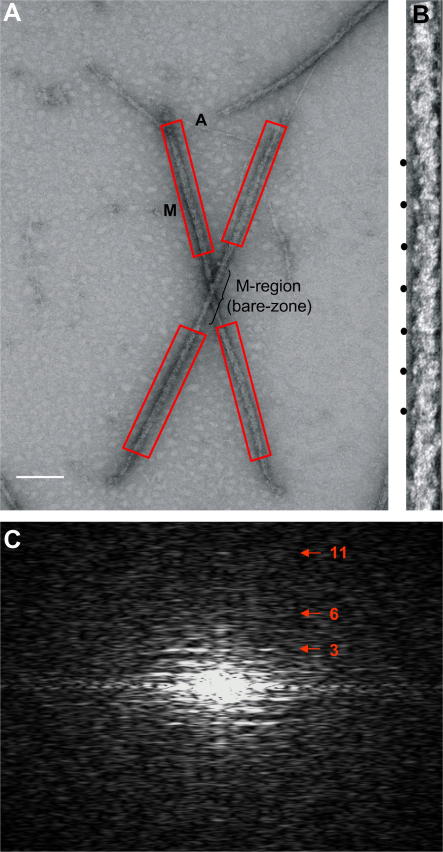
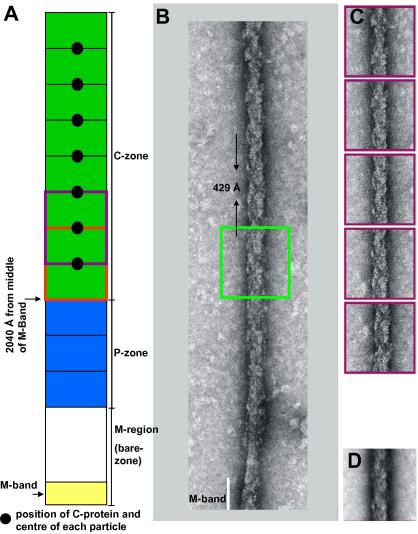
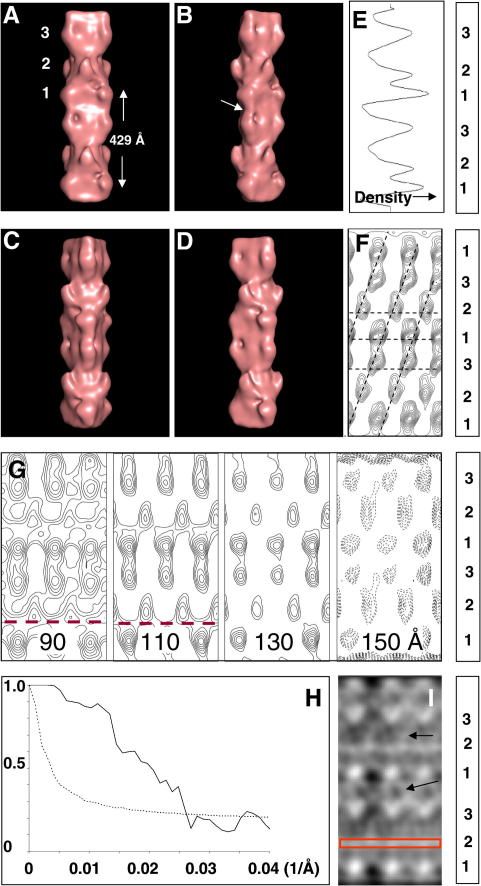
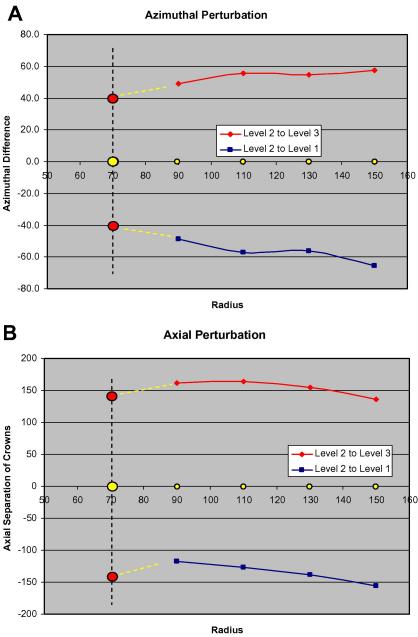
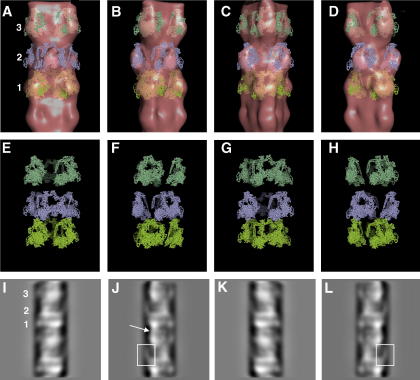
A number of cardiac myopathies (e.g. familial hypertrophic cardiomyopathy and dilated cardiomyopathy) are linked to mutations in cardiac muscle myosin filament proteins, including myosin and myosin binding protein C (MyBP-C). To understand the myopathies it is necessary to know the normal 3D structure of these filaments. We have carried out 3D single particle analysis of electron micrograph images of negatively stained isolated myosin filaments from rabbit cardiac muscle. Single filament images were aligned and divided into segments about 2x430A long, each of which was treated as an independent 'particle'. The resulting 40A resolution 3D reconstruction showed both axial and azimuthal (no radial) myosin head perturbations within the 430A repeat, with successive crown rotations of approximately 60 degrees , 60 degrees and 0 degrees , rather than the regular 40 degrees for an unperturbed helix. However, it is shown that the projecting density peaks appear to start at low radius from origins closer to those expected for an unperturbed helical filament, and that the azimuthal perturbation especially increases with radius. The head arrangements in rabbit cardiac myosin filaments are very similar to those in fish skeletal muscle myosin filaments, suggesting a possible general structural theme for myosin filaments in all vertebrate striated muscles (skeletal and cardiac).
Figures
References
-
- Aksoy M.O., Williams D., Sharkey E.M., Hartshorne D.J. A relationship between Ca2+ sensitivity and phosphorylation of gizzard actomyosin. Biochem. Biophys. Res. Commun. 1976;69:35–41. - PubMed
-
- AL-Khayat H.A., Morris E.P., Squire J.M. Single particle analysis: a new approach to solving the 3D structure of myosin filaments. Review. J. Muscle Res. Cell Motil. 2004;25(8):635–644. - PubMed
-
- AL-Khayat H.A., Squire J.M. Refined structure of bony fish muscle myosin filaments from low-angle X-ray diffraction Data. J. Struct. Biol. 2006;155:218–229. - PubMed
-
- AL-Khayat H.A., Morris E.P., Kensler R.W., Squire J.M. 3D structure of fish muscle myosin filaments by single particle analysis. J. Struct. Biol. 2006;155:202–217. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
