

The synthetic triterpenoid CDDO-methyl ester modulates microglial activities, inhibits TNF production, and provides dopaminergic neuroprotection
- PMID: 18474101
- PMCID: PMC2396606
- DOI: 10.1186/1742-2094-5-14
The synthetic triterpenoid CDDO-methyl ester modulates microglial activities, inhibits TNF production, and provides dopaminergic neuroprotection
Abstract
Background: Recent animal and human studies implicate chronic activation of microglia in the progressive loss of CNS neurons. The inflammatory mechanisms that have neurotoxic effects and contribute to neurodegeneration need to be elucidated and specifically targeted without interfering with the neuroprotective effects of glial activities. Synthetic triterpenoid analogs of oleanolic acid, such as methyl-2-cyano-3,12-dioxooleana-1,9-dien-28-oate (CDDO-Me, RTA 402) have potent anti-proliferative and differentiating effects on tumor cells, and anti-inflammatory activities on activated macrophages. We hypothesized that CDDO-Me may be able to suppress neurotoxic microglial activities while enhancing those that promote neuronal survival. Therefore, the aims of our study were to identify specific microglial activities modulated by CDDO-Me in vitro, and to determine the extent to which this modulation affords neuroprotection against inflammatory stimuli.
Methods: We tested the synthetic triterpenoid methyl-2-cyano-3,12-dioxooleana-1,9-dien-28-oate (CDDO-Me, RTA 402) in various in vitro assays using the murine BV2 microglia cell line, mouse primary microglia, or mouse primary peritoneal macrophages to investigate its effects on proliferation, inflammatory gene expression, cytokine secretion, and phagocytosis. The antioxidant and neuroprotective effects of CDDO-Me were also investigated in primary neuron/glia cultures from rat basal forebrain or ventral midbrain.
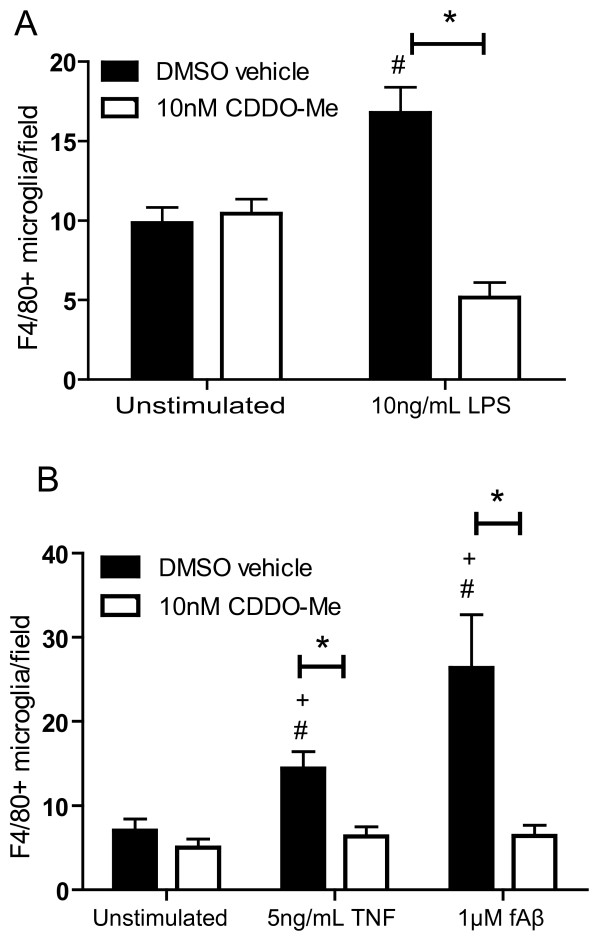
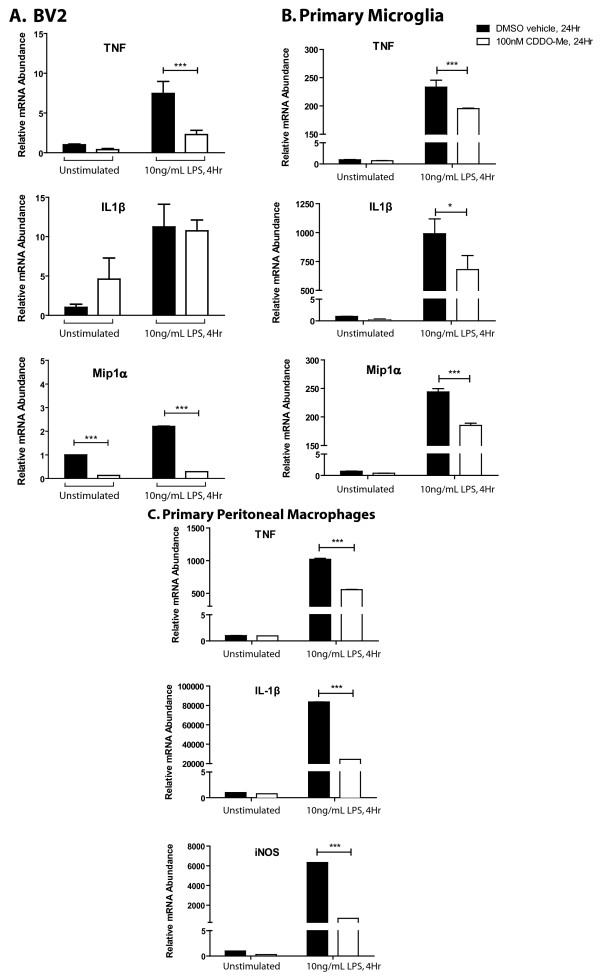
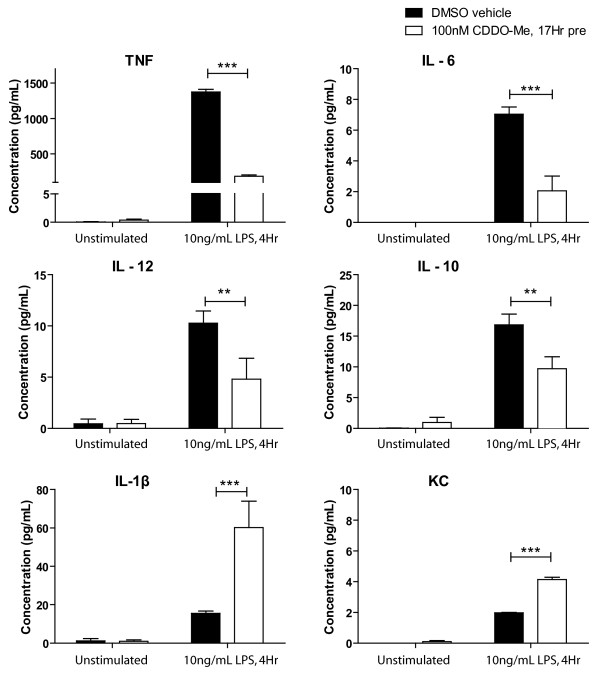
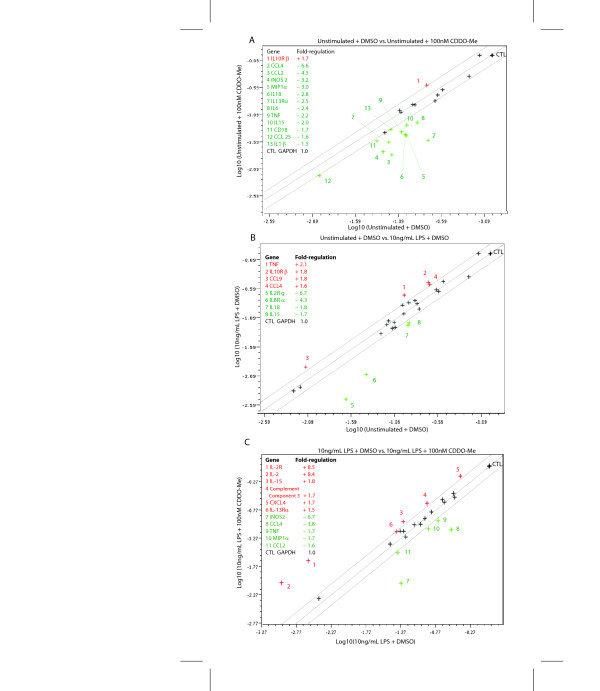
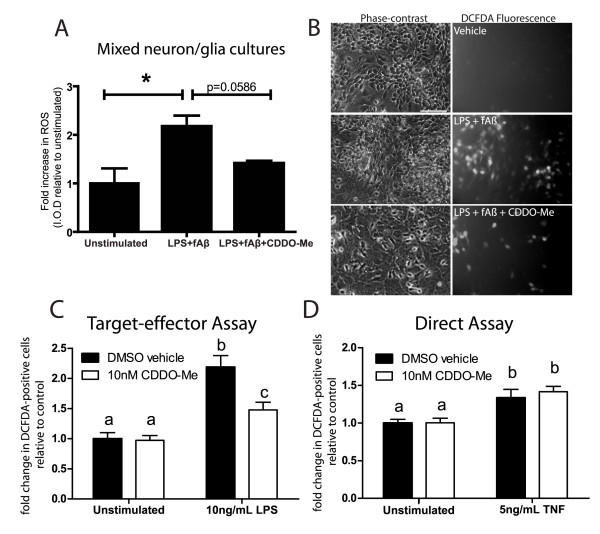
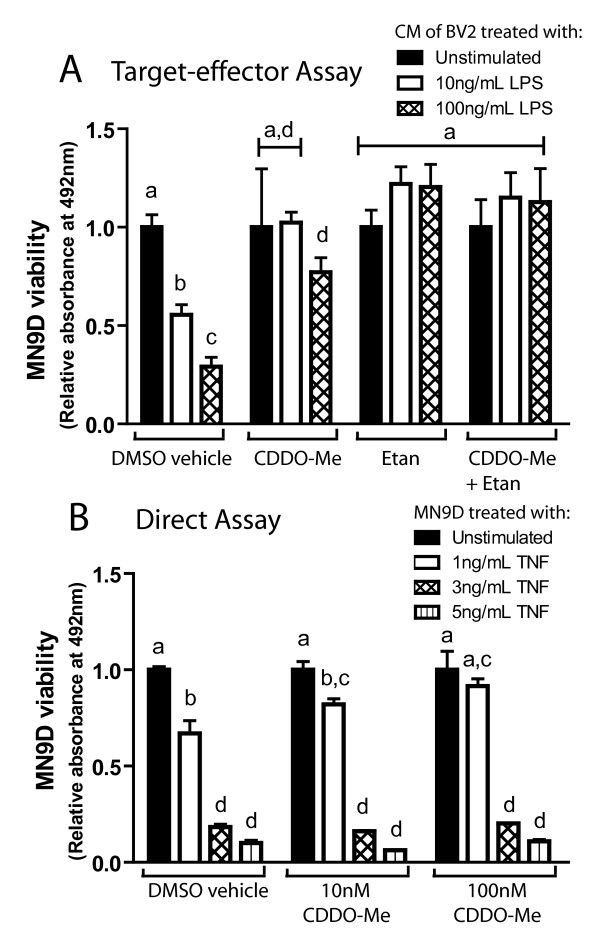
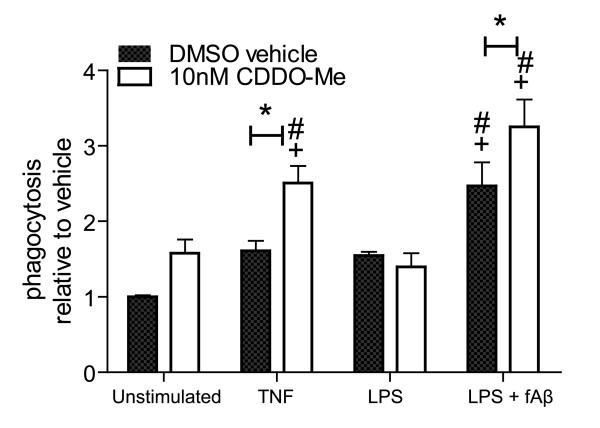
Results: We found that at low nanomolar concentrations, treatment of rat primary mesencephalon neuron/glia cultures with CDDO-Me resulted in attenuated LPS-, TNF- or fibrillar amyloid beta 1-42 (A beta 1-42) peptide-induced increases in reactive microglia and inflammatory gene expression without an overall effect on cell viability. In functional assays CDDO-Me blocked death in the dopaminergic neuron-like cell line MN9D induced by conditioned media (CM) of LPS-stimulated BV2 microglia, but did not block cell death induced by addition of TNF to MN9D cells, suggesting that dopaminergic neuroprotection by CDDO-Me involved inhibition of microglial-derived cytokine production and not direct inhibition of TNF-dependent pro-apoptotic pathways. Multiplexed immunoassays of CM from LPS-stimulated microglia confirmed that CDDO-Me-treated BV2 cells produced decreased levels of specific subsets of cytokines, in particular TNF. Lastly, CDDO-Me enhanced phagocytic activity of BV2 cells in a stimulus-specific manner but inhibited generation of reactive oxygen species (ROS) in mixed neuron/glia basal forebrain cultures and dopaminergic cells.
Conclusion: The neuroimmune modulatory properties of CDDO-Me indicate that this potent antioxidant and anti-inflammatory compound may have therapeutic potential to modify the course of neurodegenerative diseases characterized by chronic neuroinflammation and amyloid deposition. The extent to which synthetic triterpenoids afford therapeutic benefit in animal models of Parkinson's and Alzheimer's disease deserves further investigation.
Figures
References
-
- Honda T, Rounds BV, Bore L, Finlay HJ, Favaloro FG, Jr, Suh N, Wang Y, Sporn MB, Gribble GW. Synthetic oleanane and ursane triterpenoids with modified rings A and C: a series of highly active inhibitors of nitric oxide production in mouse macrophages. J Med Chem. 2000;43:4233–4246. doi: 10.1021/jm0002230. - DOI - PubMed
-
- Honda T, Rounds BV, Gribble GW, Suh N, Wang Y, Sporn MB. Design and synthesis of 2-cyano-3,12-dioxoolean-1,9-dien-28-oic acid, a novel and highly active inhibitor of nitric oxide production in mouse macrophages. Bioorg Med Chem Lett. 1998;8:2711–2714. doi: 10.1016/S0960-894X(98)00479-X. - DOI - PubMed
-
- Dinkova-Kostova AT, Liby KT, Stephenson KK, Holtzclaw WD, Gao X, Suh N, Williams C, Risingsong R, Honda T, Gribble GW, et al. Extremely potent triterpenoid inducers of the phase 2 response: correlations of protection against oxidant and inflammatory stress. Proc Natl Acad Sci USA. 2005;102:4584–4589. doi: 10.1073/pnas.0500815102. - DOI - PMC - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
