

MglA and Igl proteins contribute to the modulation of Francisella tularensis live vaccine strain-containing phagosomes in murine macrophages
- PMID: 18474647
- PMCID: PMC2493230
- DOI: 10.1128/IAI.00226-08
MglA and Igl proteins contribute to the modulation of Francisella tularensis live vaccine strain-containing phagosomes in murine macrophages
Abstract
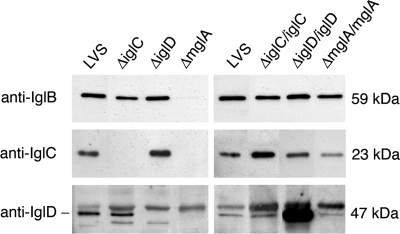
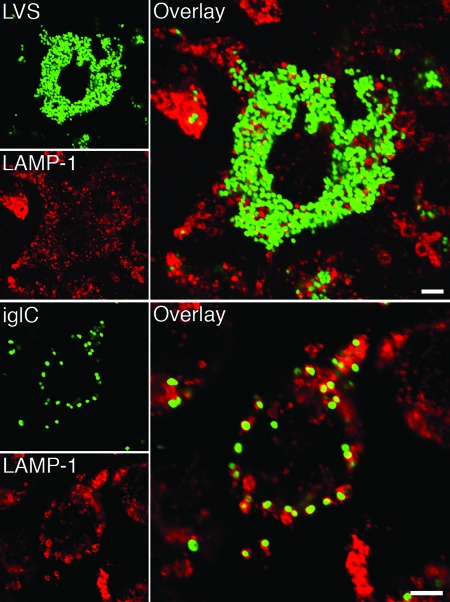
The Francisella tularensis live vaccine strain (LVS), in contrast to its iglC mutant, replicates in the cytoplasm of macrophages. We studied the outcome of infection of the murine macrophagelike cell line J774A.1 with LVS and with iglC, iglD, and mglA mutants, the latter of which is deficient in a global regulator. Compared to LVS, all of the mutants showed impaired intracellular replication up to 72 h, and the number of the mglA mutant bacteria even decreased. Colocalization with LAMP-1 was significantly increased for all mutants compared to LVS, indicating an impaired ability to escape into the cytoplasm. A lysosomal acidity-dependent dye accumulated in approximately 40% of the vacuoles containing mutant bacteria but not at all in vacuoles containing LVS. Preactivation of the macrophages with gamma interferon inhibited the intracellular growth of all strains and significantly increased acidification of phagosomes containing the mutants, but it only slightly increased the LAMP-1 colocalization. The intracellular replication and phagosomal escape of the iglC and iglD mutants were restored by complementation in trans. In conclusion, the IglC, IglD, and MglA proteins each directly or indirectly critically contribute to the virulence of F. tularensis LVS, including its intracellular replication, cytoplasmic escape, and inhibition of acidification of the phagosomes.
Figures
References
-
- Anthony, L. S., P. J. Morrissey, and F. E. Nano. 1992. Growth inhibition of Francisella tularensis live vaccine strain by IFN-gamma-activated macrophages is mediated by reactive nitrogen intermediates derived from l-arginine metabolism. J. Immunol. 1481829-1834. - PubMed
-
- Brumell, J. H., and S. Grinstein. 2004. Salmonella redirects phagosomal maturation. Curr. Opin. Microbiol. 778-84. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
