

Sensory signaling-dependent remodeling of olfactory cilia architecture in C. elegans
- PMID: 18477458
- PMCID: PMC2442577
- DOI: 10.1016/j.devcel.2008.03.002
Sensory signaling-dependent remodeling of olfactory cilia architecture in C. elegans
Abstract
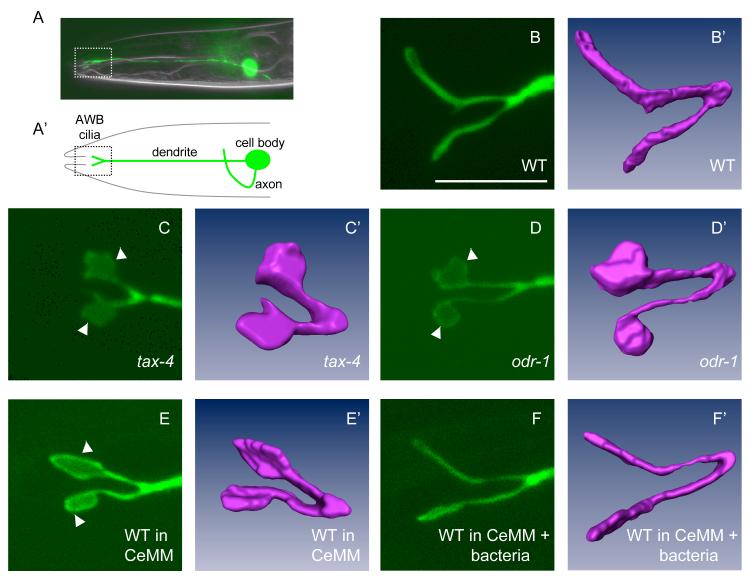
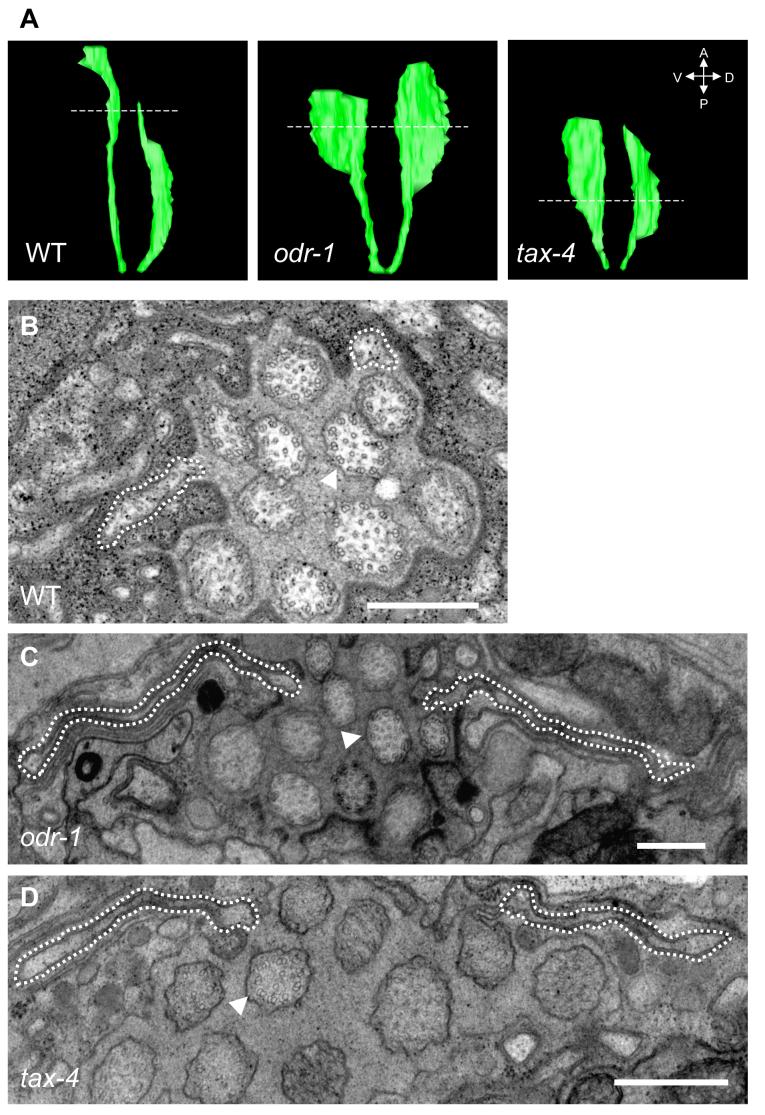
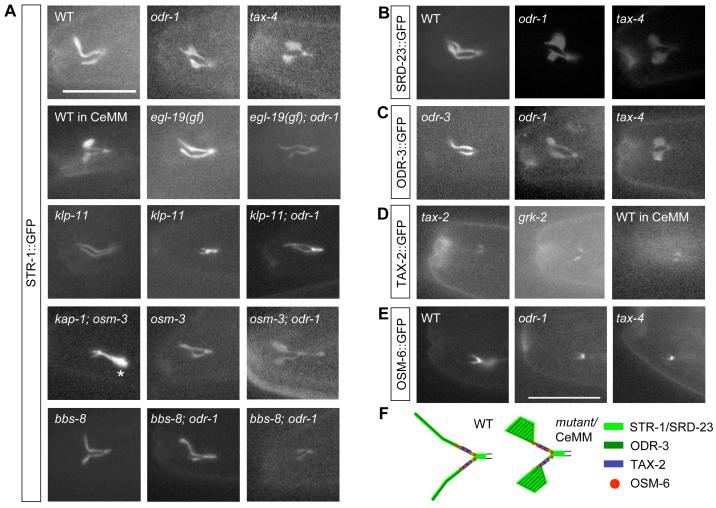
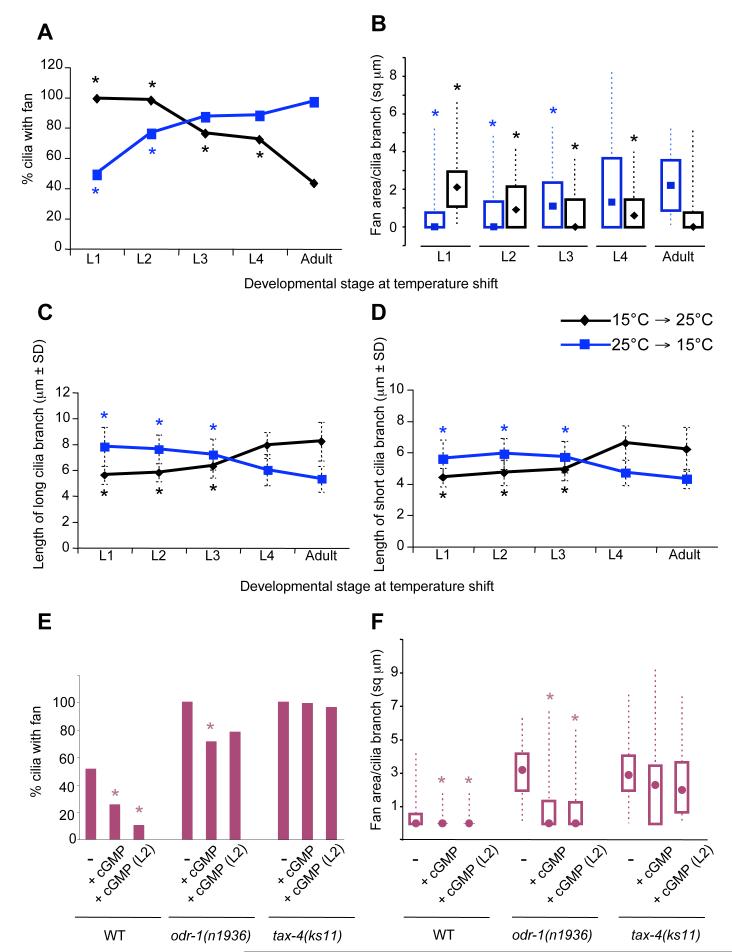
Nonmotile primary cilia are sensory organelles composed of a microtubular axoneme and a surrounding membrane sheath that houses signaling molecules. Optimal cellular function requires the precise regulation of axoneme assembly, membrane biogenesis, and signaling protein targeting and localization via as yet poorly understood mechanisms. Here, we show that sensory signaling is required to maintain the architecture of the specialized AWB olfactory neuron cilia in C. elegans. Decreased sensory signaling results in alteration of axoneme length and expansion of a membraneous structure, thereby altering the topological distribution of a subset of ciliary transmembrane signaling molecules. Signaling-regulated alteration of ciliary structures can be bypassed by modulation of intracellular cGMP or calcium levels and requires kinesin-II-driven intraflagellar transport (IFT), as well as BBS- and RAB8-related proteins. Our results suggest that compensatory mechanisms in response to altered levels of sensory activity modulate AWB cilia architecture, revealing remarkable plasticity in the regulation of cilia structure.
Figures
Comment in
-
A cilium is not a cilium is not a cilium: signaling contributes to ciliary morphological diversity.Dev Cell. 2008 May;14(5):635-6. doi: 10.1016/j.devcel.2008.04.011. Dev Cell. 2008. PMID: 18477443
References
-
- Alvarez VA, Sabatini BL. Anatomical and physiological plasticity of dendritic spines. Annu. Rev. Neurosci. 2007;30:79–97. - PubMed
-
- Ansley SJ, Badano JL, Blacque OE, Hill J, Hoskins BE, Leitch CC, Kim JC, Ross AJ, Eichers ER, Teslovich TM, et al. Basal body dysfunction is a likely cause of pleiotropic Bardet-Biedl syndrome. Nature. 2003;425:628–633. - PubMed
-
- Bargmann CI, Hartwieg E, Horvitz HR. Odorant-selective genes and neurons mediate olfaction in C. elegans. Cell. 1993;74:515–527. - PubMed
-
- Bargmann CI, Horvitz HR. Chemosensory neurons with overlapping functions direct chemotaxis to multiple chemicals in C. elegans. Neuron. 1991;7:729–742. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
