

Growth/differentiation factor 3 signals through ALK7 and regulates accumulation of adipose tissue and diet-induced obesity
- PMID: 18480259
- PMCID: PMC2438236
- DOI: 10.1073/pnas.0800272105
Growth/differentiation factor 3 signals through ALK7 and regulates accumulation of adipose tissue and diet-induced obesity
Abstract
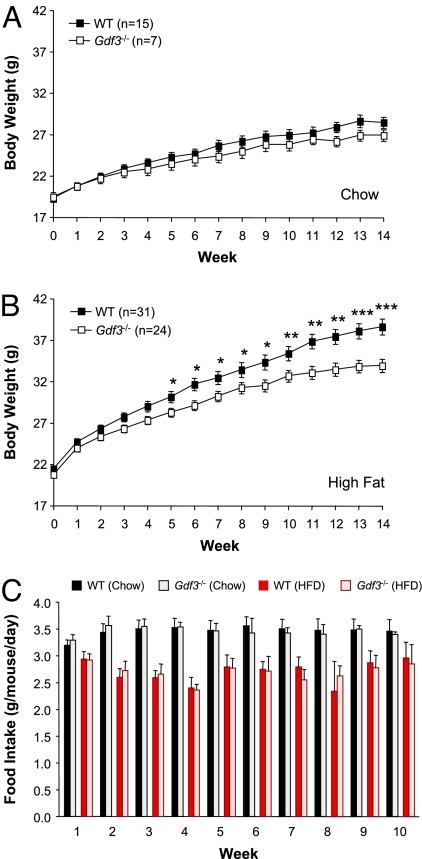
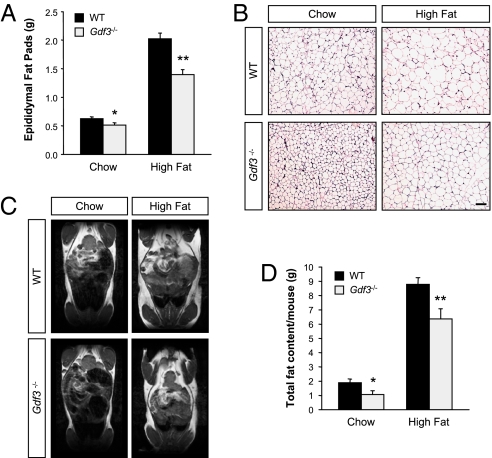
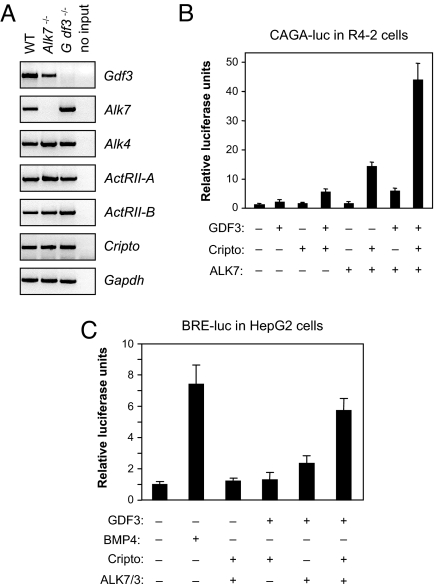
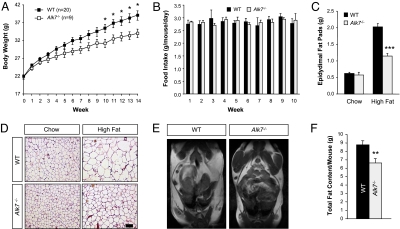
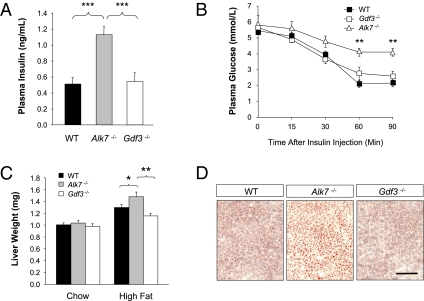
Growth/differentiation factor 3 (GDF3) is highly expressed in adipose tissue, and previous overexpression experiments in mice have suggested that it may act as an adipogenic factor under conditions of high lipid load. GDF3 has been shown to signal via the activin receptor ALK4 during embryogenesis, but functional receptors in adipose tissue are unknown. In this study, we show that Gdf3(-/-) mutant mice accumulate less adipose tissue than WT animals and show partial resistance to high-fat diet-induced obesity despite similar food intake. We also demonstrate that GDF3 can signal via the ALK4-homolog ALK7 and the coreceptor Cripto, both of which are expressed in adipose tissue. In agreement with a role for ALK7 in GDF3 signaling in vivo, mutant mice lacking ALK7 also showed reduced fat accumulation and partial resistance to diet-induced obesity. We propose that GDF3 regulates adipose-tissue homeostasis and energy balance under nutrient overload in part by signaling through the ALK7 receptor.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
