

Live cell dynamics of promyelocytic leukemia nuclear bodies upon entry into and exit from mitosis
- PMID: 18480407
- PMCID: PMC2441680
- DOI: 10.1091/mbc.e08-01-0035
Live cell dynamics of promyelocytic leukemia nuclear bodies upon entry into and exit from mitosis
Abstract
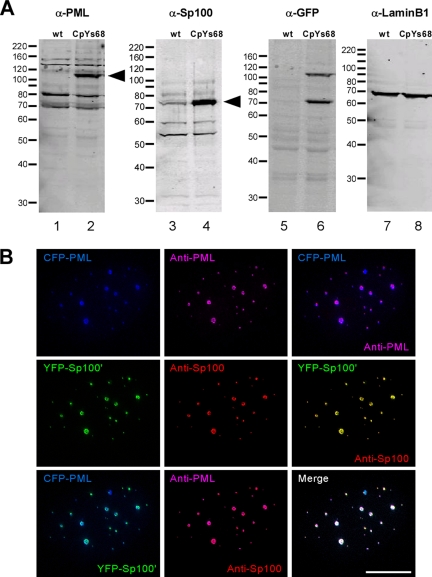
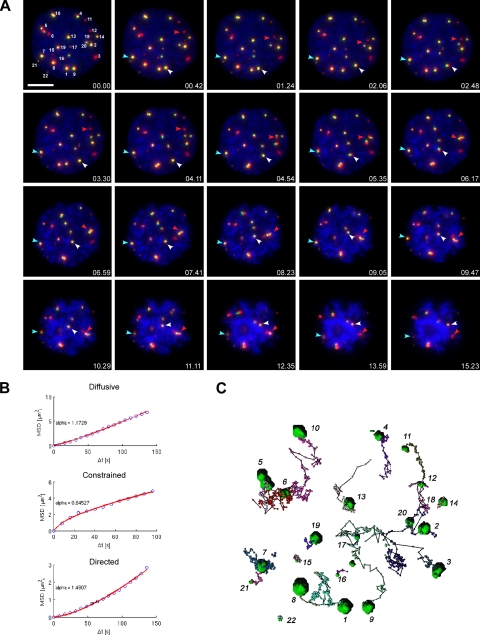
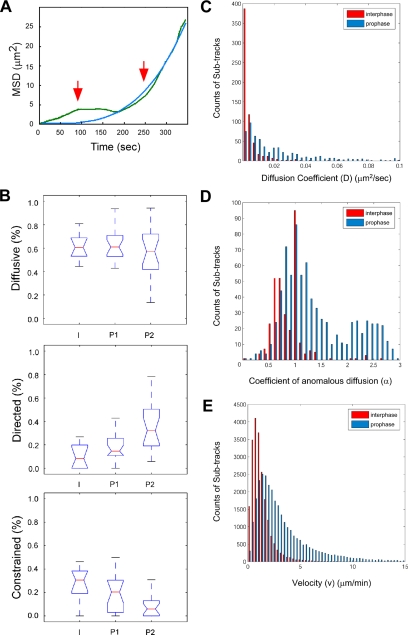
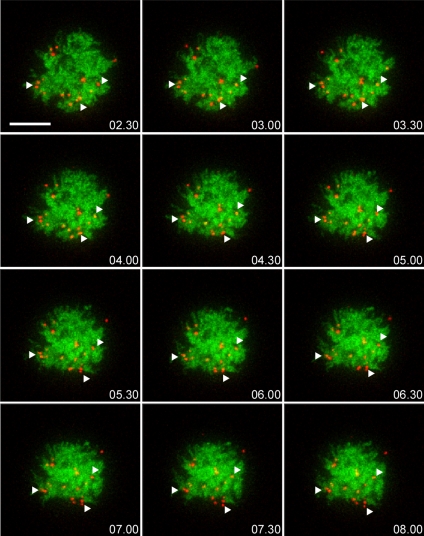
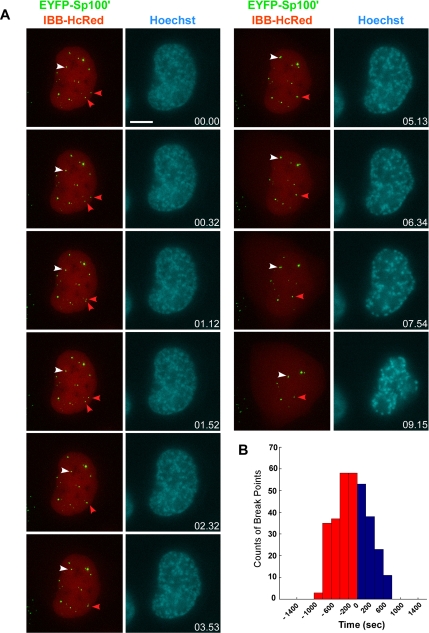
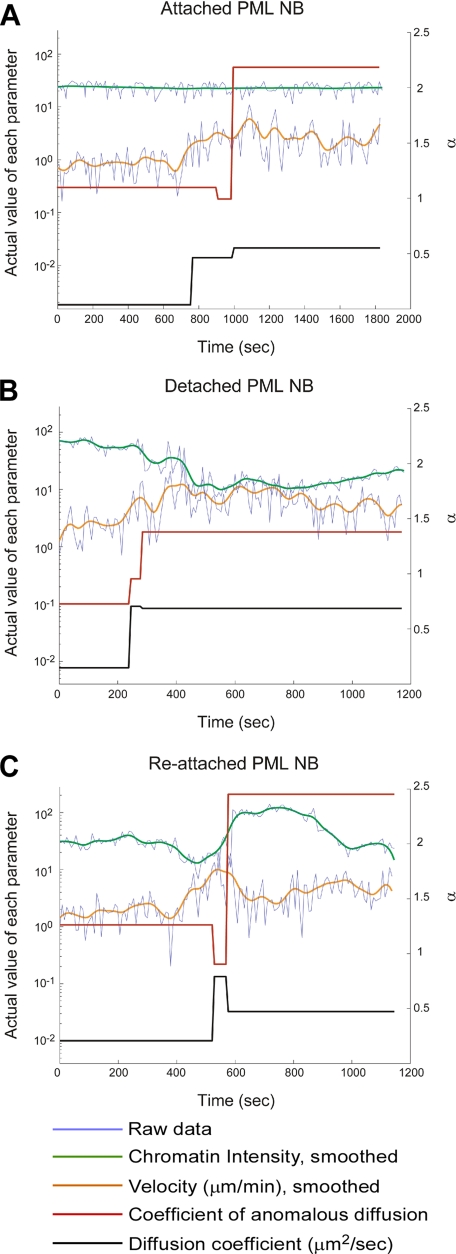
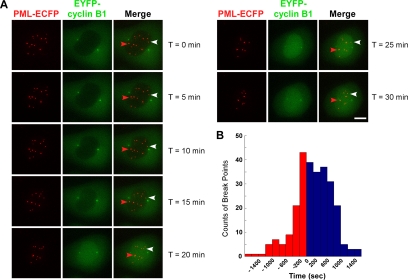
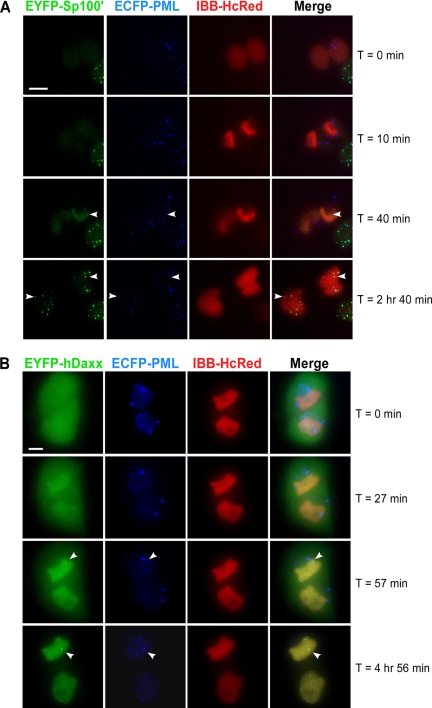
Promyelocytic leukemia nuclear bodies (PML NBs) have been proposed to be involved in tumor suppression, viral defense, DNA repair, and/or transcriptional regulation. To study the dynamics of PML NBs during mitosis, we developed several U2OS cell lines stably coexpressing PML-enhanced cyan fluorescent protein with other individual marker proteins. Using three-dimensional time-lapse live cell imaging and four-dimensional particle tracking, we quantitatively demonstrated that PML NBs exhibit a high percentage of directed movement when cells progressed from prophase to prometaphase. The timing of this increased dynamic movement occurred just before or upon nuclear entry of cyclin B1, but before nuclear envelope breakdown. Our data suggest that entry into prophase leads to a loss of tethering between regions of chromatin and PML NBs, resulting in their increased dynamics. On exit from mitosis, Sp100 and Fas death domain-associated protein (Daxx) entered the daughter nuclei after a functional nuclear membrane was reformed. However, the recruitment of these proteins to PML NBs was delayed and correlated with the timing of de novo PML NB formation. Together, these results provide insight into the dynamic changes associated with PML NBs during mitosis.
Figures
References
-
- Albiez H., et al. Chromatin domains and the interchromatin compartment form structurally defined and functionally interacting nuclear networks. Chromosome Res. 2006;14:707–733. - PubMed
-
- Asakawa Y., Gotoh E. A method for detecting sister chromatid exchanges using prematurely condensed chromosomes and immunogold-silver staining. Mutagenesis. 1997;12:175–177. - PubMed
-
- Bassermann F., Peschel C., Duyster J. Mitotic entry: a matter of oscillating destruction. Cell Cycle. 2005;4:1515–1517. - PubMed
-
- Bernardi R., Pandolfi P. P. Structure, dynamics and functions of promyelocytic leukaemia nuclear bodies. Nat. Rev. Mol. Cell Biol. 2007;8:1006–1016. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
