

Extreme dependence of gH and gL expression on ORF57 and association with highly unusual codon usage in rhesus monkey rhadinovirus
- PMID: 18480451
- PMCID: PMC2446953
- DOI: 10.1128/JVI.00564-08
Extreme dependence of gH and gL expression on ORF57 and association with highly unusual codon usage in rhesus monkey rhadinovirus
Abstract
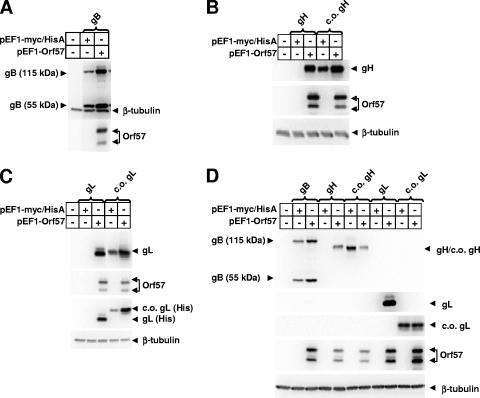
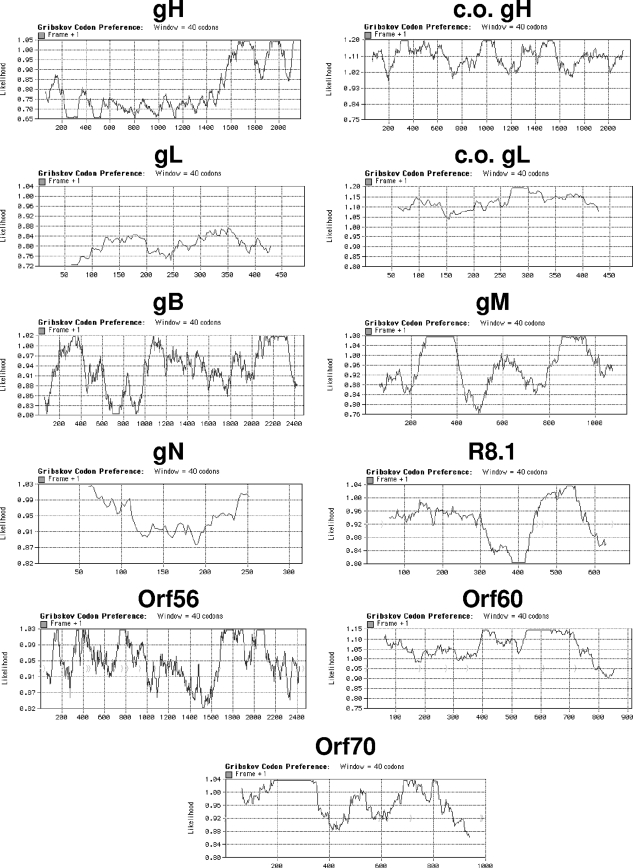
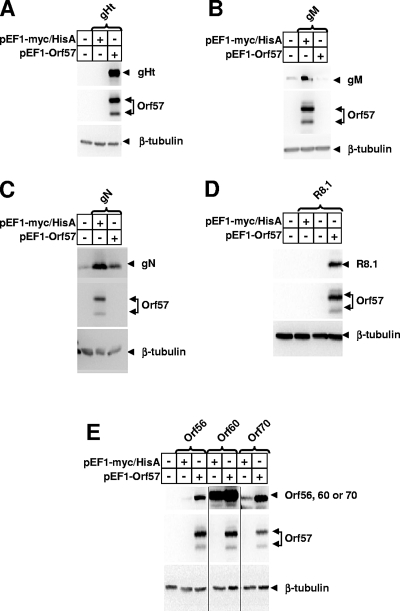
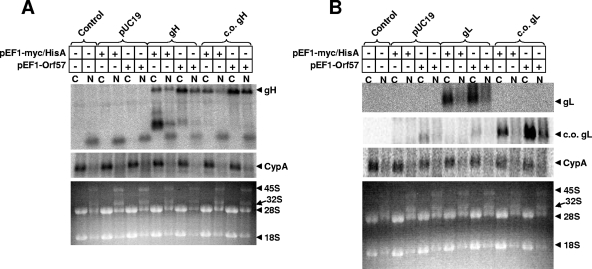
Standard vectors for high-level expression elicited undetectable levels of the gH and gL glycoproteins of rhesus monkey rhadinovirus (RRV) following transient-transfection assays under a variety of conditions. These same vectors and conditions yielded high levels of RRV gB expression. Unlike other genes of RRV, both the gH and gL genes were noted to have a highly aberrant, suboptimal codon usage. High levels of RRV gH and gL expression were achieved by two alternative means: codon optimization or coexpression of RRV ORF57. The failure of gH and gL to be expressed in the absence of ORF57 and in the absence of codon optimization could not be explained by the failure of RNA to egress from the nucleus. Rather, the defect in gH and gL expression appeared to be cytoplasmic in nature. It is not clear at the present time whether the aberrant codon usage for gH and gL of RRV is an intentional regulatory strategy used by the virus or whether it is driven by some external force, such as intrinsic immunity. In any event, our results indicate that the need of ORF57 for gH and gL expression can be circumvented by codon optimization, that RRV ORF57 acts principally to allow translation of gH and gL RNA in the cytoplasm, and that this activity of ORF57 is related in some way to the aberrant codon usage of the gH and gL RNAs.
Figures
References
-
- Bishop, K. N., R. K. Holmes, A. M. Sheehy, and M. H. Malim. 2004. APOBEC-mediated editing of viral RNA. Science 305645. - PubMed
-
- Boyne, J. R., and A. Whitehouse. 2006. Gamma-2 herpes virus post-transcriptional gene regulation. Clin. Microbiol. Infect. 12110-117. - PubMed
-
- Bulmer, M. 1987. Coevolution of codon usage and transfer RNA abundance. Nature 325728-730. - PubMed
-
- Carbone, A., A. Zinovyev, and F. Kepes. 2003. Codon adaptation index as a measure of dominating codon bias. Bioinformatics 192005-2015. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
