

Loss of heparan sulfate glycosaminoglycan assembly in podocytes does not lead to proteinuria
- PMID: 18480751
- PMCID: PMC8092444
- DOI: 10.1038/ki.2008.159
Loss of heparan sulfate glycosaminoglycan assembly in podocytes does not lead to proteinuria
Abstract
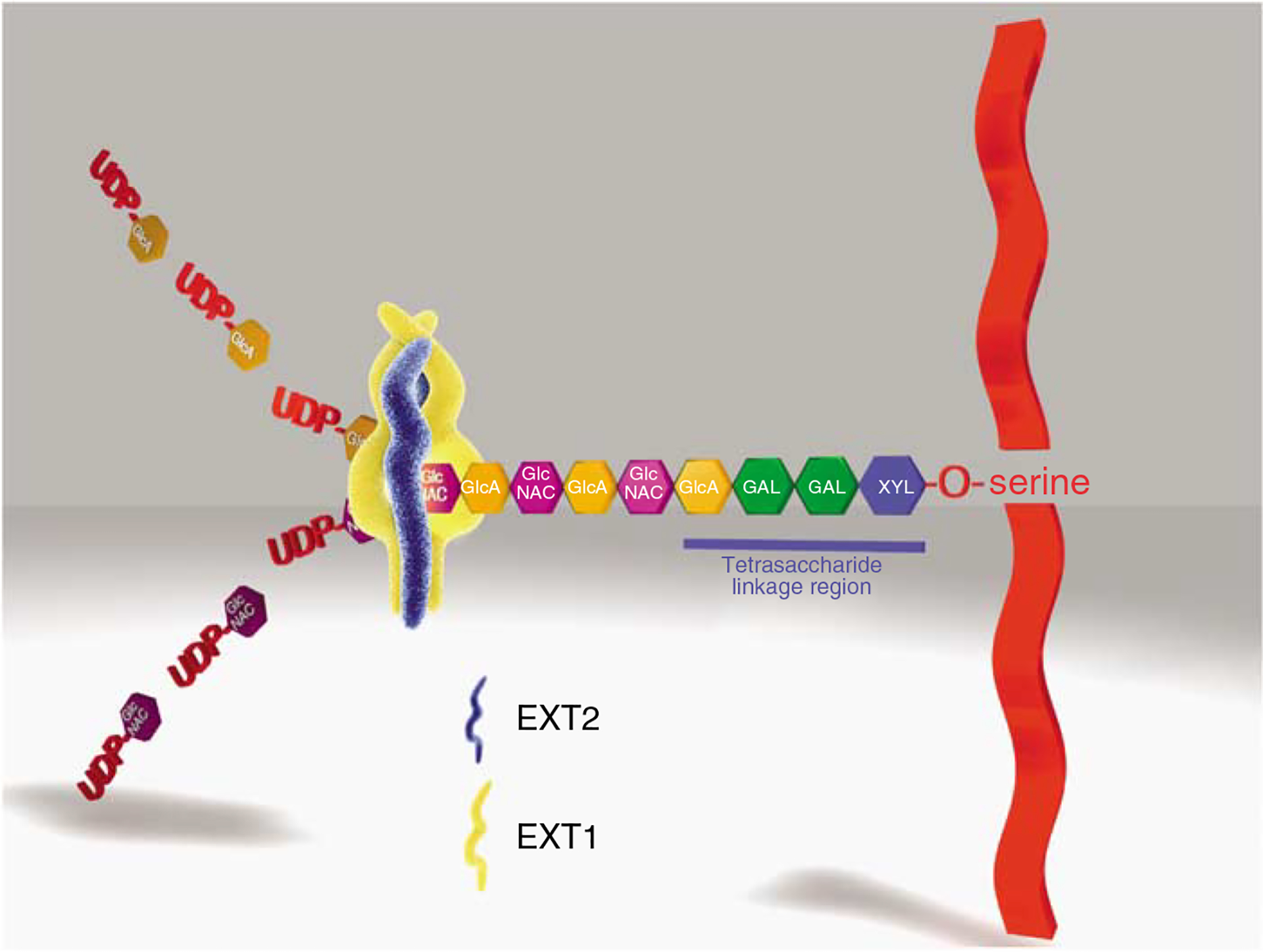
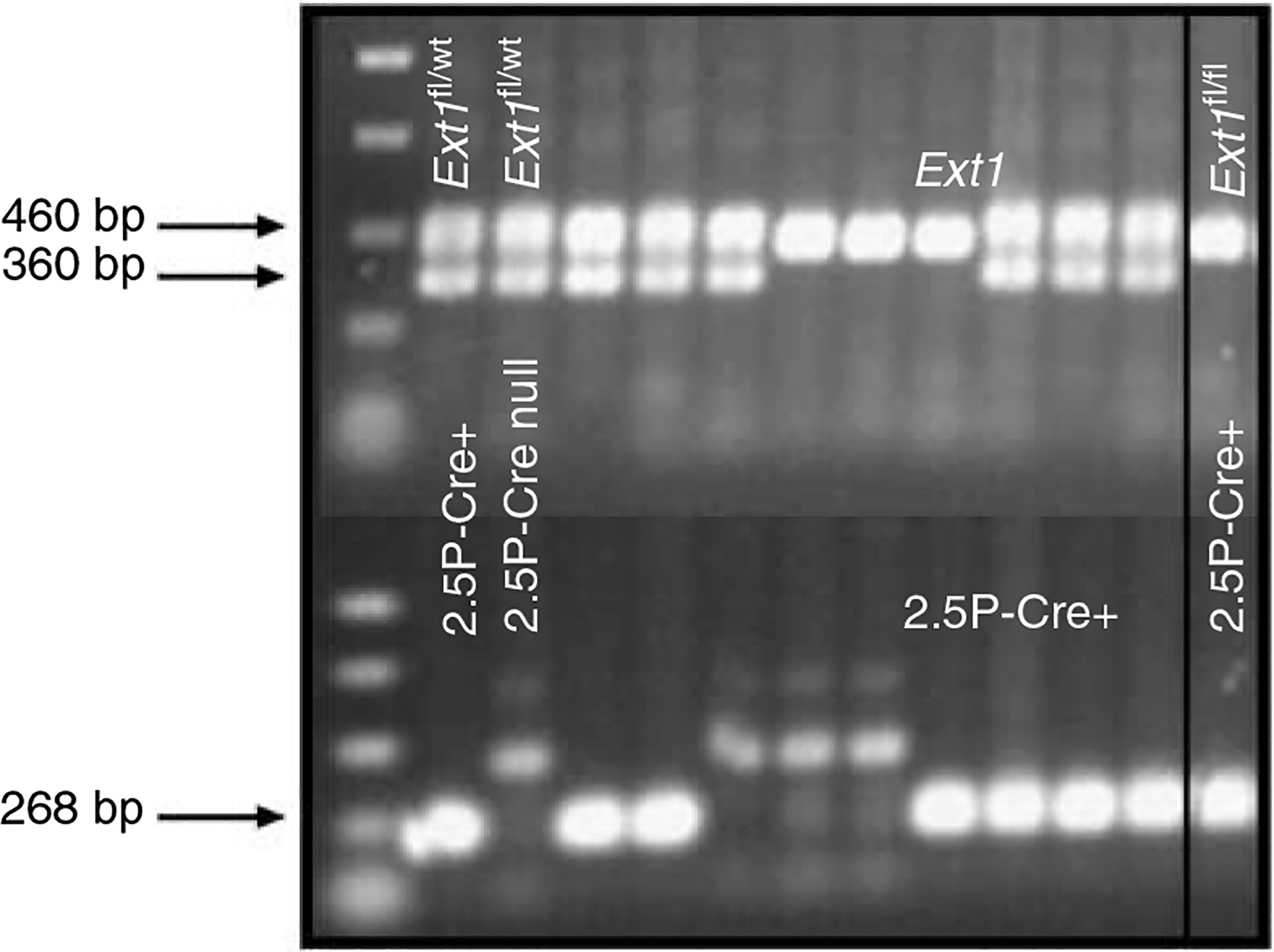
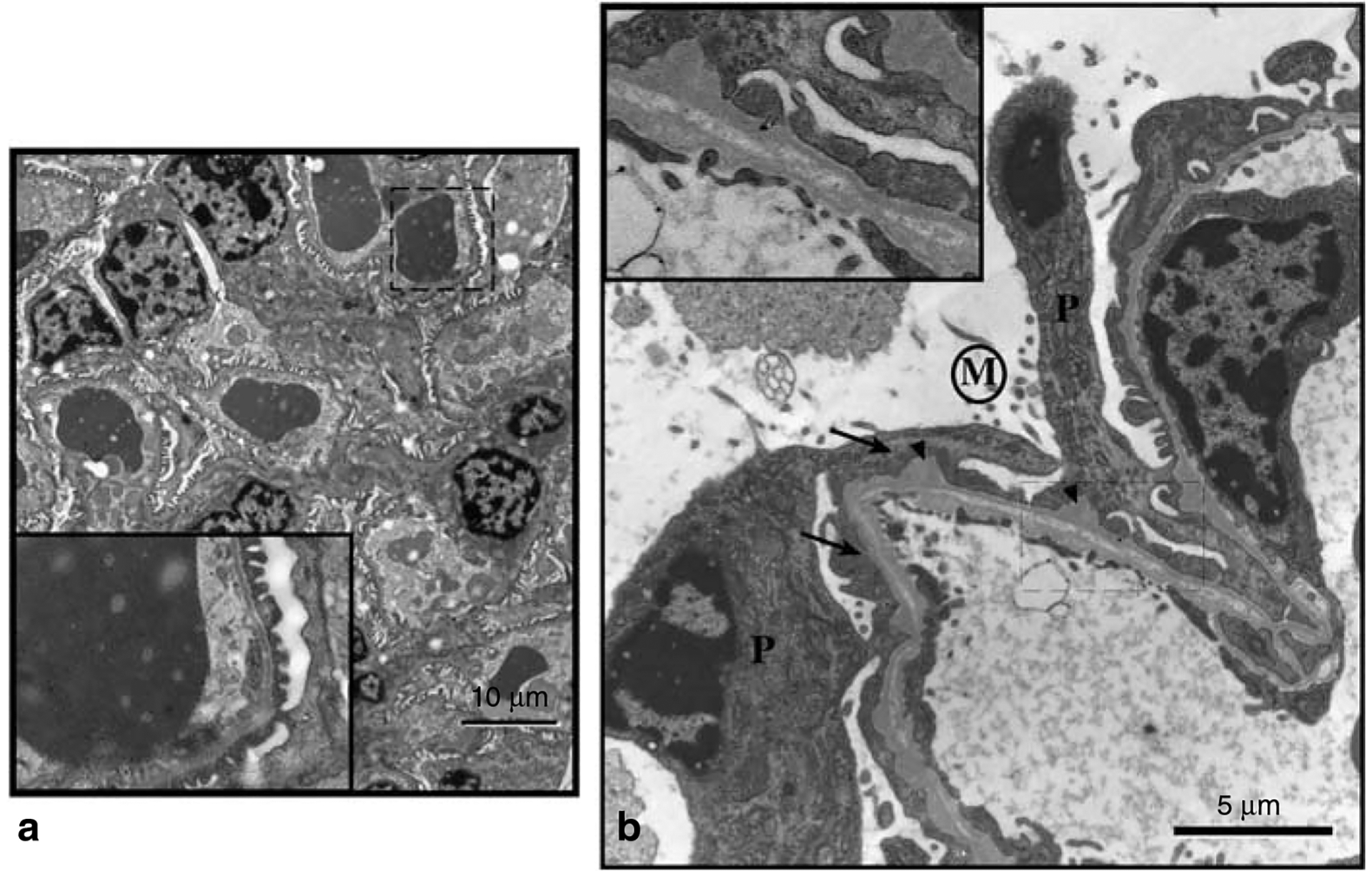
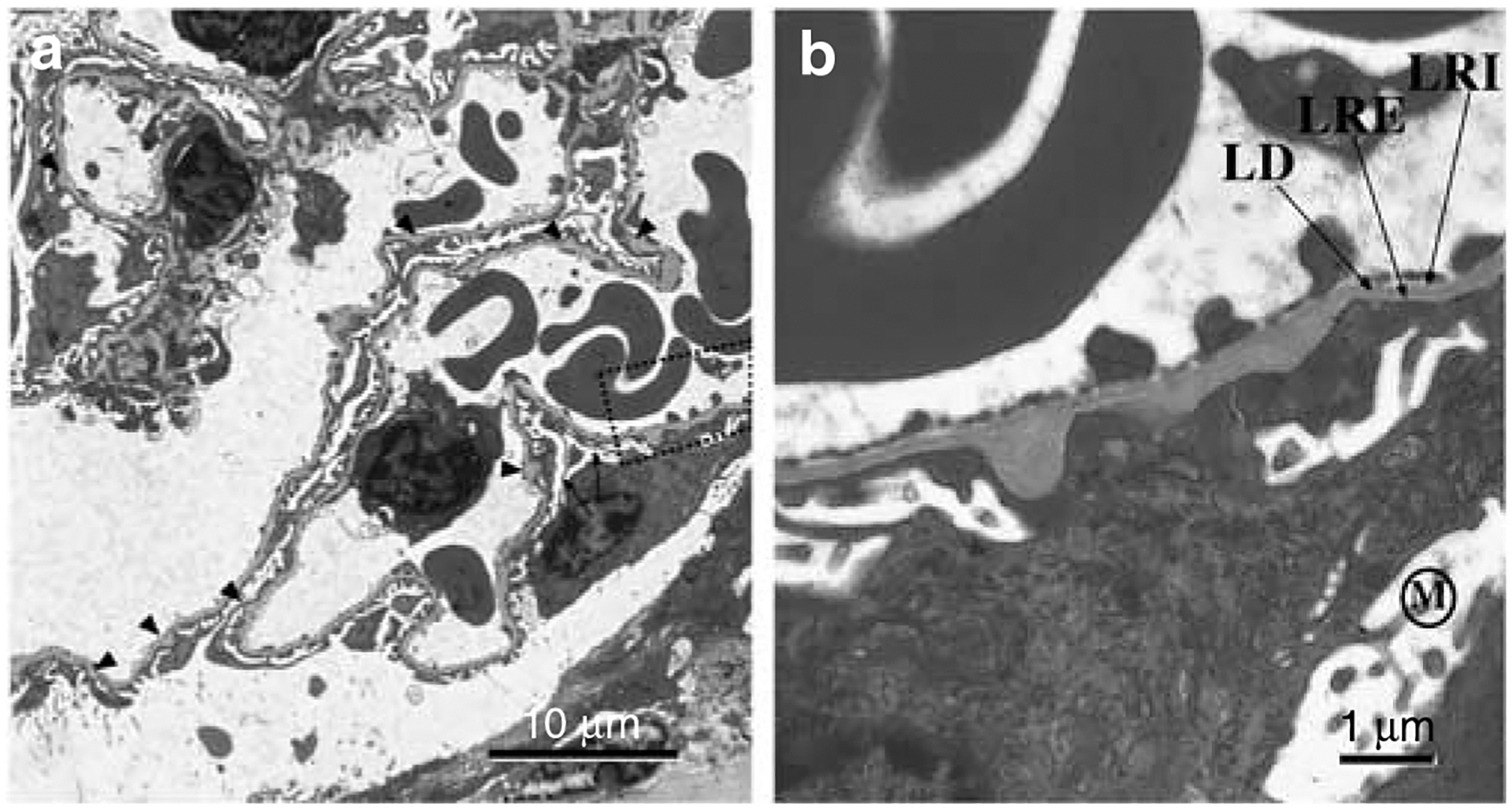
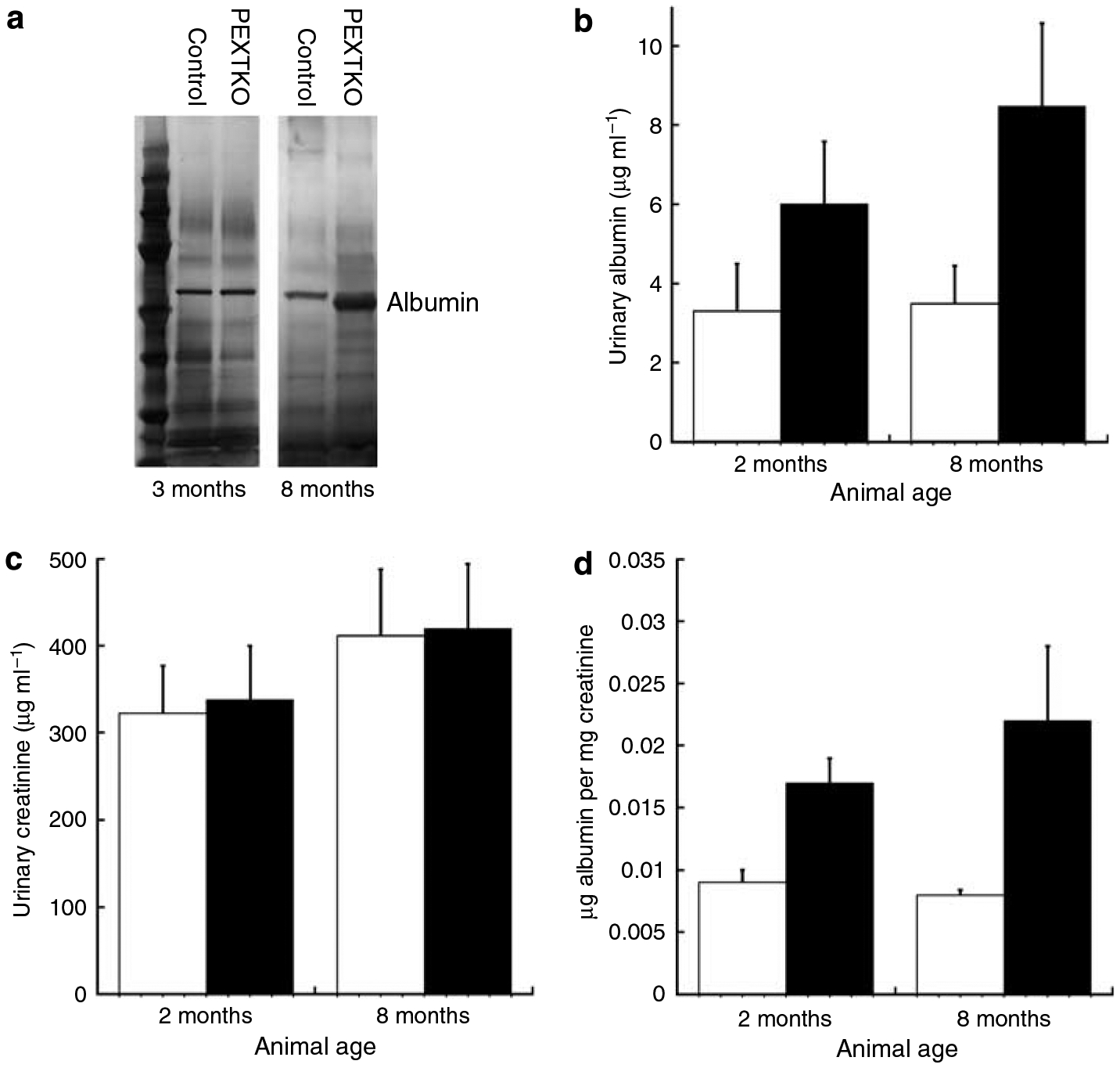
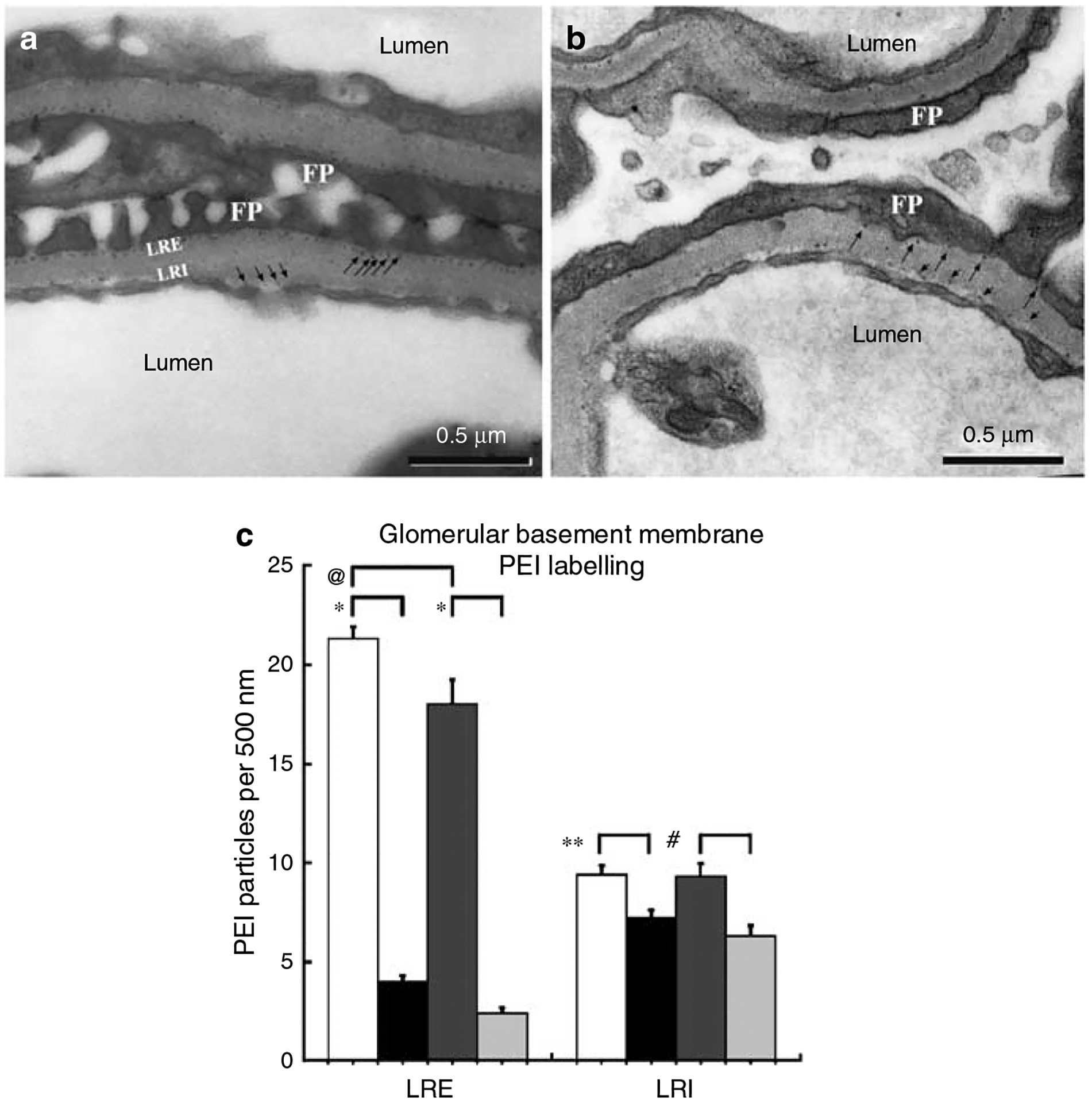
Podocytes synthesize the majority of the glomerular basement membrane components with some contribution from the glomerular capillary endothelial cells. The anionic charge of heparan sulfate proteoglycans is conferred by covalently attached heparan sulfate glycosaminoglycans and these are thought to provide critical charge selectivity to the glomerular basement membrane for ultrafiltration. One key component in herparan sulfate glycosaminoglycan assembly is the Ext1 gene product encoding a subunit of heparan sulfate co-polymerase. Here we knocked out Ext1 gene expression in podocytes halting polymerization of heparin sulfate glycosaminoglycans on the proteoglycan core proteins secreted by podocytes. Glomerular development occurred normally in these knockout animals but changes in podocyte morphology, such as foot process effacement, were seen as early as 1 month after birth. Immunohistochemical analysis showed a significant decrease in heparan sulfate glycosaminoglycans confirmed by ultrastructural studies using polyethyleneimine staining. Despite podocyte abnormalities and loss of heparan sulfate glycosaminoglycans, severe albuminuria did not develop in the knockout mice. We show that the presence of podocyte-secreted heparan sulfate glycosaminoglycans is not absolutely necessary to limit albuminuria suggesting the existence of other mechanisms that limit albuminuria. Heparan sulfate glycosaminoglycans appear to have functions that control podocyte behavior rather than be primarily an ultrafiltration barrier.
Conflict of interest statement
DISCLOSURE
All the authors declared no competing interests.
Figures
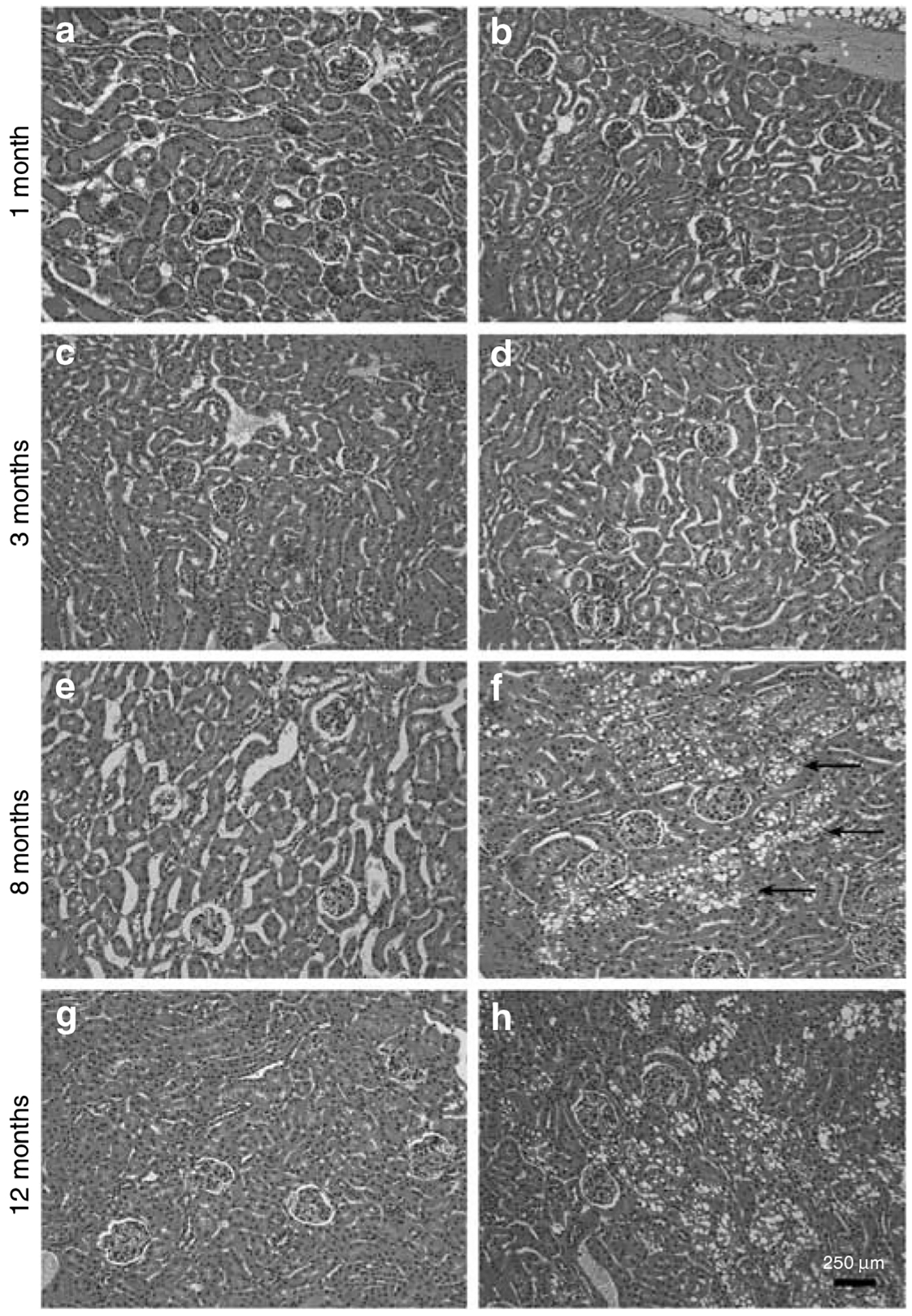
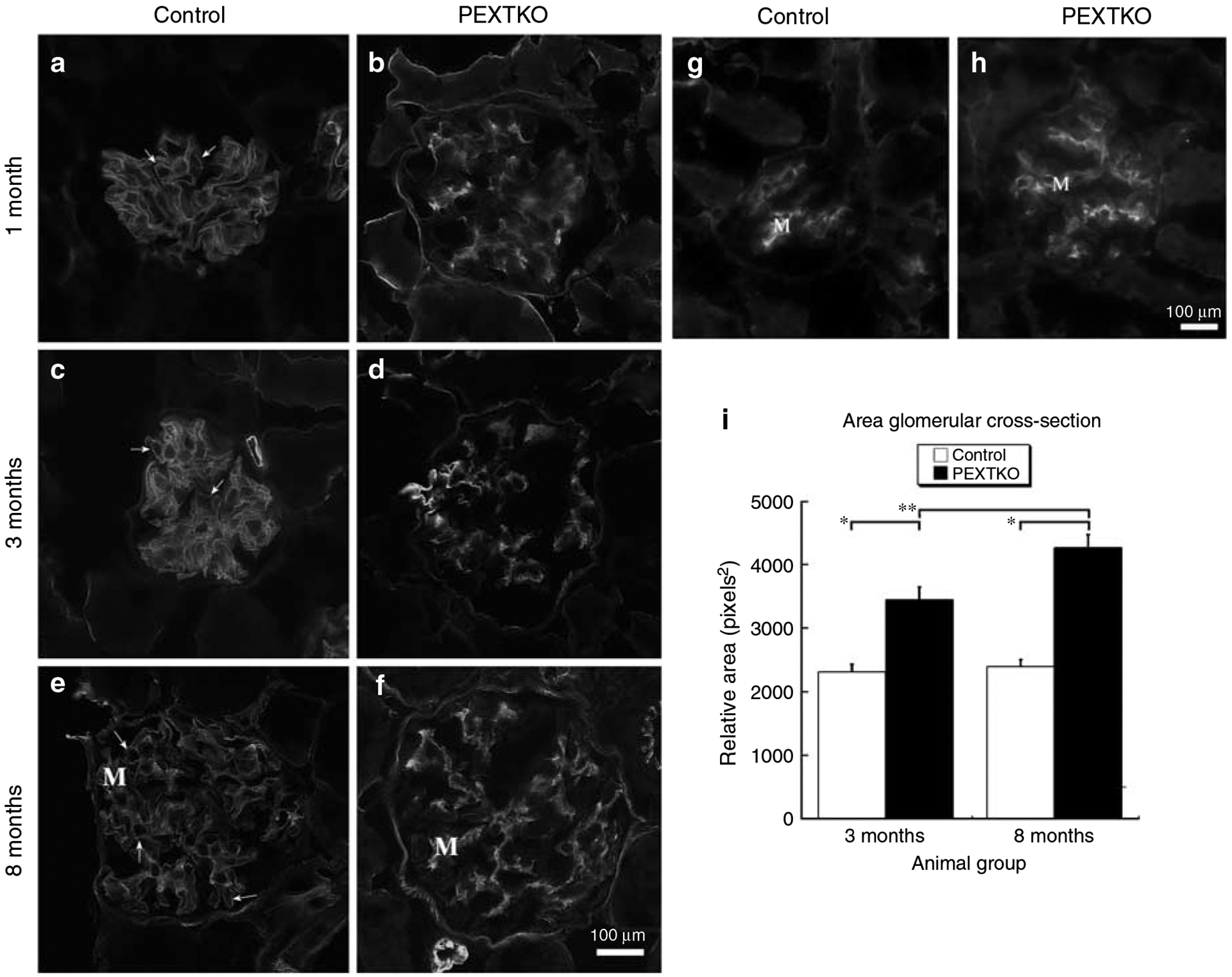
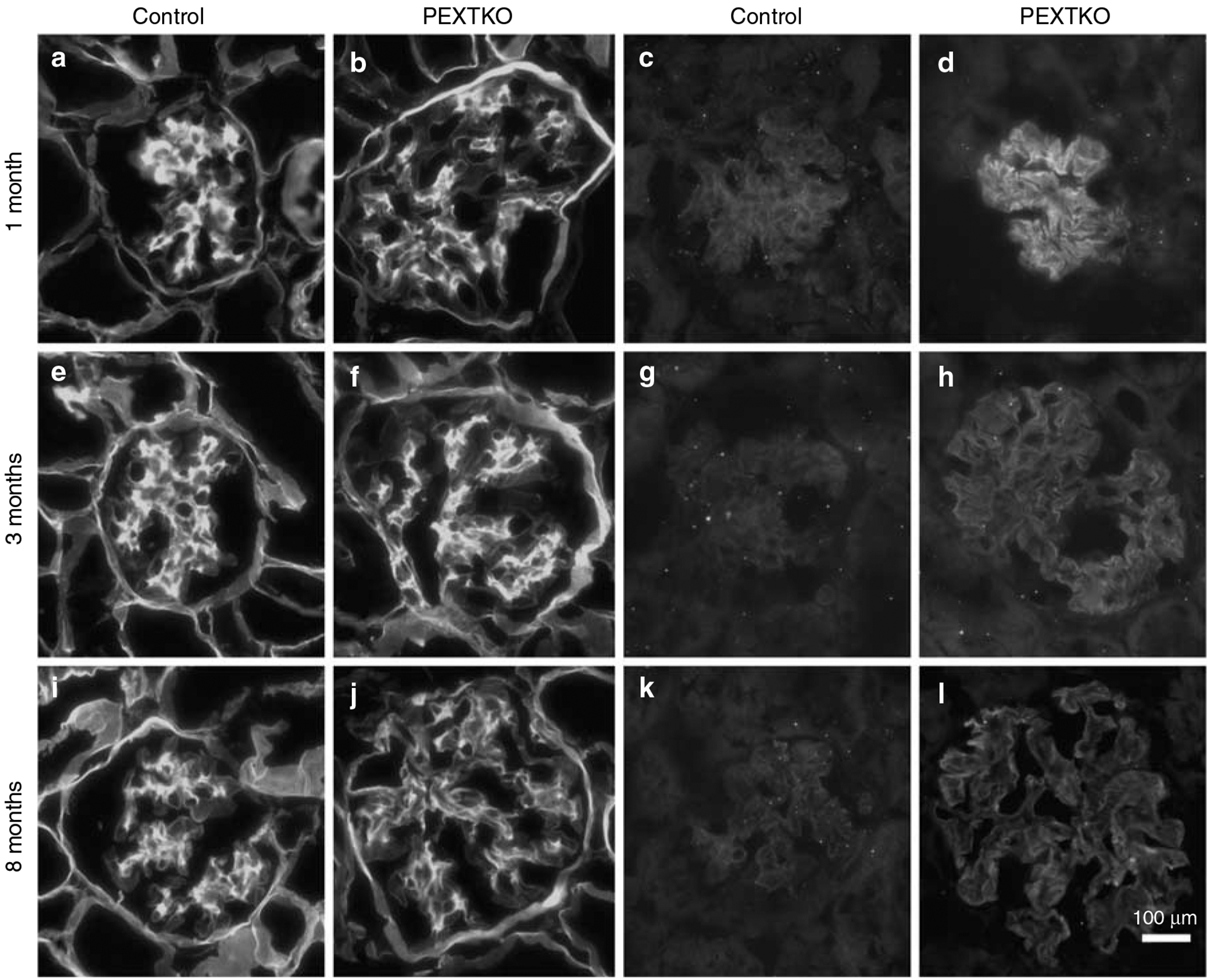
 control 2 months,
control 2 months,  PEXTKO 2 months,
PEXTKO 2 months,  control 8 months,
control 8 months,  PEXTKO 8 months.
PEXTKO 8 months.Comment in
-
Glomerular filtration: the charge debate charges ahead.Kidney Int. 2008 Aug;74(3):259-61. doi: 10.1038/ki.2008.260. Kidney Int. 2008. PMID: 18626493
References
-
- Iozzo RV, Murdoch AD. Proteoglycans of the extracellular environment: clues from the gene and protein side offer novel perspectives in molecular diversity and function. FASEB J 1996; 10: 598–614. - PubMed
-
- Esko JD, Selleck SB. Order out of chaos: assembly of ligand binding sites in heparan sulfate. Annu Rev Biochem 2002; 71: 435–471. - PubMed
-
- Cattaruzza S, Perris R. Proteoglycan control of cell movement during wound healing and cancer spreading. Matrix Biol 2005; 24: 400–417. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
