

Potential molecular mechanism for c-Src kinase-mediated regulation of intestinal cell migration
- PMID: 18482983
- PMCID: PMC2504885
- DOI: 10.1074/jbc.M801319200
Potential molecular mechanism for c-Src kinase-mediated regulation of intestinal cell migration
Abstract
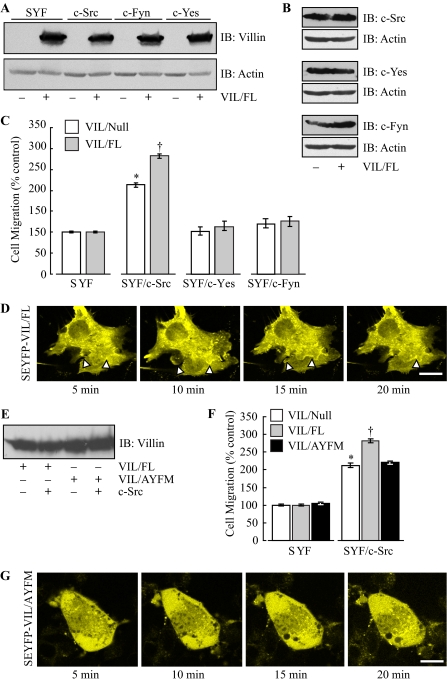
The ubiquitously expressed Src tyrosine kinases (c-Src, c-Yes, and c-Fyn) regulate intestinal cell growth and differentiation. Src activity is also elevated in the majority of malignant and premalignant tumors of the colon. The development of fibroblasts with the three ubiquitously expressed kinases deleted (SYF cells) has identified the role of Src proteins in the regulation of actin dynamics associated with increased cell migration and invasion. Despite this, unexpectedly nothing is known about the role of the individual Src kinases on intestinal cell cytoskeleton and/or cell migration. We have previously reported that villin, an epithelial cell-specific actin-modifying protein that regulates actin reorganization, cell morphology, cell migration, cell invasion, and apoptosis, is tyrosine-phosphorylated. In this report using the SYF cells reconstituted individually with c-Src, c-Yes, c-Fyn, and wild type or phosphorylation site mutants of villin, we demonstrate for the first time the absolute requirement for c-Src in villin-induced regulation of cell migration. The other major finding of our study is that contrary to previous reports, the nonreceptor tyrosine kinase, Jak3 (Janus kinase 3), does not regulate phosphorylation of villin or villin-induced cell migration and is, in fact, not expressed in intestinal epithelial cells. Further, we identify SHP-2 and PTP-PEST (protein-tyrosine phosphatase proline-, glutamate-, serine-, and threonine-rich sequence) as negative regulators of c-Src kinase and demonstrate a new function for these phosphatases in intestinal cell migration. Together, these data suggest that in colorectal carcinogenesis, elevation of c-Src or down-regulation of SHP-2 and/or PTP-PEST may promote cancer metastases and invasion by regulating villin-induced cell migration and cell invasion.
Figures






Similar articles
-
Regulation of cell motility by tyrosine phosphorylated villin.Mol Biol Cell. 2004 Nov;15(11):4807-17. doi: 10.1091/mbc.e04-05-0431. Epub 2004 Sep 1. Mol Biol Cell. 2004. PMID: 15342783 Free PMC article.
-
Interaction of Pyk2 and PTP-PEST with leupaxin in prostate cancer cells.Am J Physiol Cell Physiol. 2007 Jun;292(6):C2288-96. doi: 10.1152/ajpcell.00503.2006. Epub 2007 Feb 28. Am J Physiol Cell Physiol. 2007. PMID: 17329398
-
Regulation of phospholipase C-gamma(1) by the actin-regulatory protein villin.Am J Physiol Cell Physiol. 2001 Sep;281(3):C1046-58. doi: 10.1152/ajpcell.2001.281.3.C1046. Am J Physiol Cell Physiol. 2001. PMID: 11502583
-
Regulation of tumor cell migration by protein tyrosine phosphatase (PTP)-proline-, glutamate-, serine-,and threonine-rich sequence (PEST).Chin J Cancer. 2013 Feb;32(2):75-83. doi: 10.5732/cjc.012.10084. Epub 2012 Dec 7. Chin J Cancer. 2013. PMID: 23237212 Free PMC article. Review.
-
The potent role of Src kinase-regulating glucose metabolism in cancer.Biochem Pharmacol. 2022 Dec;206:115333. doi: 10.1016/j.bcp.2022.115333. Epub 2022 Oct 29. Biochem Pharmacol. 2022. PMID: 36404485 Review.
Cited by
-
Maresin-2 promotes mucosal repair and has therapeutic potential when encapsulated in thermostable nanoparticles.Proc Natl Acad Sci U S A. 2023 Jan 24;120(4):e2218162120. doi: 10.1073/pnas.2218162120. Epub 2023 Jan 20. Proc Natl Acad Sci U S A. 2023. PMID: 36669099 Free PMC article.
-
Cross talk between receptor guanylyl cyclase C and c-src tyrosine kinase regulates colon cancer cell cytostasis.Mol Cell Biol. 2009 Oct;29(19):5277-89. doi: 10.1128/MCB.00001-09. Epub 2009 Jul 20. Mol Cell Biol. 2009. PMID: 19620276 Free PMC article.
-
Actin reorganization as the molecular basis for the regulation of apoptosis in gastrointestinal epithelial cells.Cell Death Differ. 2012 Sep;19(9):1514-24. doi: 10.1038/cdd.2012.28. Epub 2012 Mar 16. Cell Death Differ. 2012. PMID: 22421965 Free PMC article.
-
Genetic variability in EGFR, Src and HER2 and risk of colorectal adenoma and cancer.Int J Mol Epidemiol Genet. 2011;2(4):300-15. Epub 2011 Dec 3. Int J Mol Epidemiol Genet. 2011. PMID: 22199994 Free PMC article.
-
PTP-PEST targets a novel tyrosine site in p120 catenin to control epithelial cell motility and Rho GTPase activity.J Cell Sci. 2014 Feb 1;127(Pt 3):497-508. doi: 10.1242/jcs.120154. Epub 2013 Nov 27. J Cell Sci. 2014. PMID: 24284071 Free PMC article.
References
-
- Cartwright, C. A., Mamajiwalla, S., Skolnick, S. A., Eckhart, W., and Burgess, D. R. (1993) Oncogene 8 1033-1039 - PubMed
-
- Martin, G. S. (2001) Nat. Rev. Mol. Cell Biol. 2 467-475 - PubMed
-
- Irby, R. B., Mao, W., Coppola, D., Kang, J., Loubeau, J. M., Trudeau, W., Karl, R., Fujita, D. J., Jove, R., and Yeatman, T. J. (1999) Nat. Genet. 21 187-190 - PubMed
-
- Aligayer, H., Boyd, D. D., Heiss, M. M., Abdalla, E. K., Curley, S. A., and Gallick, G. E. (2002) Cancer 94 344-351 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
