

Opposite effects of dihydrosphingosine 1-phosphate and sphingosine 1-phosphate on transforming growth factor-beta/Smad signaling are mediated through the PTEN/PPM1A-dependent pathway
- PMID: 18482992
- PMCID: PMC2443671
- DOI: 10.1074/jbc.M802417200
Opposite effects of dihydrosphingosine 1-phosphate and sphingosine 1-phosphate on transforming growth factor-beta/Smad signaling are mediated through the PTEN/PPM1A-dependent pathway
Abstract
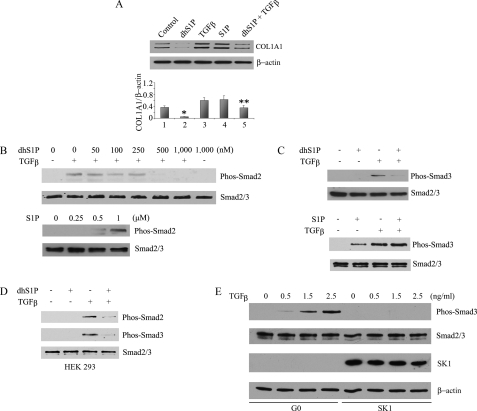
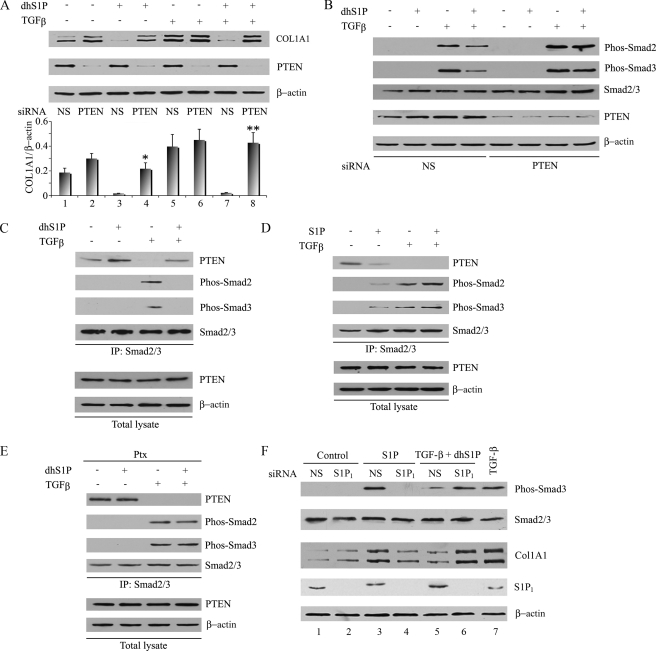
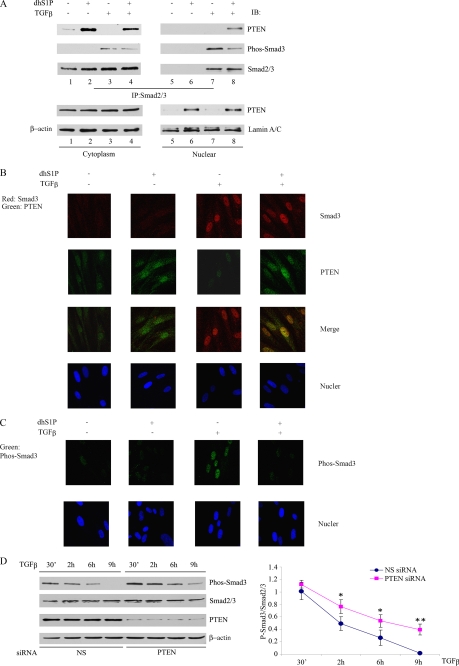
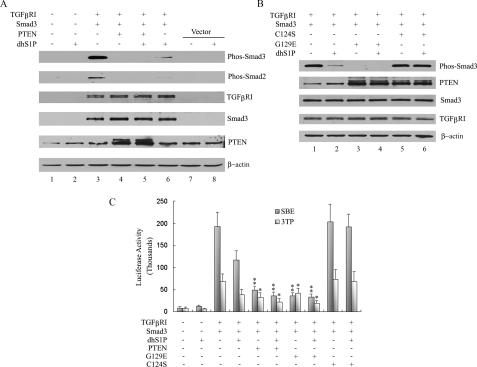
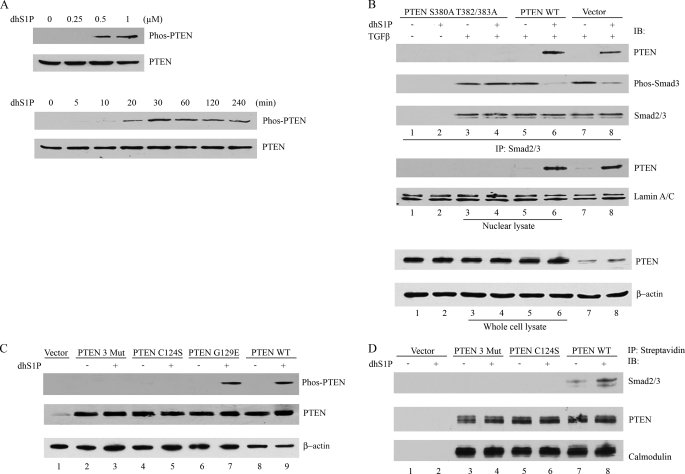
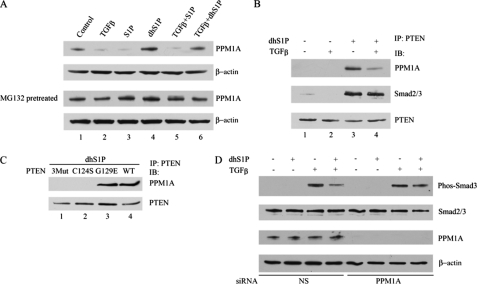
Transforming growth factor-beta (TGF-beta) is an important regulator of physiological connective tissue biosynthesis and plays a central role in pathological tissue fibrosis. Previous studies have established that a biologically active lipid mediator, sphingosine 1-phosphate (S1P), mimics some of the profibrotic functions of TGF-beta through cross-activation of Smad signaling. Here we report that another product of sphingosine kinase, dihydrosphingosine 1-phosphate (dhS1P), has an opposite role in the regulation of TGF-beta signaling. In contrast to S1P, dhS1P inhibits TGF-beta-induced Smad2/3 phosphorylation and up-regulation of collagen synthesis. The effects of dhS1P require a lipid phosphatase, PTEN, a key modulator of cell growth and survival. dhS1P stimulates phosphorylation of the C-terminal domain of PTEN and its subsequent translocation into the nucleus. We demonstrate a novel function of nuclear PTEN as a co-factor of the Smad2/3 phosphatase, PPM1A. Complex formation of PTEN with PPM1A does not require the lipid phosphatase activity but depends on phosphorylation of the serine/threonine residues located in the C-terminal domain of PTEN. Upon complex formation with PTEN, PPM1A is protected from degradation induced by the TGF-beta signaling. Consequently, overexpression of PTEN abrogates TGF-beta-induced Smad2/3 phosphorylation. This study establishes a novel role for nuclear PTEN in the stabilization of PPM1A. PTEN-mediated cross-talk between the sphingolipid and TGF-beta signaling pathways may play an important role in physiological and pathological TGF-beta signaling.
Figures
Similar articles
-
Dihydrosphingosine 1-phosphate has a potent antifibrotic effect in scleroderma fibroblasts via normalization of phosphatase and tensin homolog levels.Arthritis Rheum. 2010 Jul;62(7):2117-26. doi: 10.1002/art.27463. Arthritis Rheum. 2010. PMID: 20309867 Free PMC article.
-
PPM1A functions as a Smad phosphatase to terminate TGFbeta signaling.Cell. 2006 Jun 2;125(5):915-28. doi: 10.1016/j.cell.2006.03.044. Cell. 2006. PMID: 16751101 Free PMC article.
-
PPM1A dephosphorylates RanBP3 to enable efficient nuclear export of Smad2 and Smad3.EMBO Rep. 2011 Oct 28;12(11):1175-81. doi: 10.1038/embor.2011.174. EMBO Rep. 2011. PMID: 21960005 Free PMC article.
-
Noncanonical transforming growth factor beta signaling in scleroderma fibrosis.Curr Opin Rheumatol. 2009 Nov;21(6):623-9. doi: 10.1097/BOR.0b013e32833038ce. Curr Opin Rheumatol. 2009. PMID: 19713852 Free PMC article. Review.
-
A phosphatase controls the fate of receptor-regulated Smads.Cell. 2006 Jun 2;125(5):838-40. doi: 10.1016/j.cell.2006.05.015. Cell. 2006. PMID: 16751094 Review.
Cited by
-
Sphingosine 1-phosphate chemical biology.Biochim Biophys Acta. 2008 Sep;1781(9):508-12. doi: 10.1016/j.bbalip.2008.06.006. Epub 2008 Jun 25. Biochim Biophys Acta. 2008. PMID: 18638568 Free PMC article. Review.
-
Dihydrosphingosine 1-phosphate has a potent antifibrotic effect in scleroderma fibroblasts via normalization of phosphatase and tensin homolog levels.Arthritis Rheum. 2010 Jul;62(7):2117-26. doi: 10.1002/art.27463. Arthritis Rheum. 2010. PMID: 20309867 Free PMC article.
-
Endothelial Sphingosine-1-Phosphate Receptor 4 Regulates Blood-Brain Barrier Permeability and Promotes a Homeostatic Endothelial Phenotype.J Neurosci. 2022 Mar 9;42(10):1908-1929. doi: 10.1523/JNEUROSCI.0188-21.2021. Epub 2021 Dec 13. J Neurosci. 2022. PMID: 34903569 Free PMC article.
-
Protein phosphatase 1A (PPM1A) is involved in human cytotrophoblast cell invasion and migration.Histochem Cell Biol. 2009 Aug;132(2):169-79. doi: 10.1007/s00418-009-0601-5. Epub 2009 Apr 29. Histochem Cell Biol. 2009. PMID: 19404668
-
The role of dihydrosphingolipids in disease.Cell Mol Life Sci. 2019 Mar;76(6):1107-1134. doi: 10.1007/s00018-018-2984-8. Epub 2018 Dec 6. Cell Mol Life Sci. 2019. PMID: 30523364 Free PMC article. Review.
References
-
- Massague, J. (1998) Annu. Rev. Biochem. 67 753-791 - PubMed
-
- Bierie, B., and Moses, H. L. (2006) Nat. Rev. Cancer 6 506-520 - PubMed
-
- Bhowmick, N. A., Chytil, A., Plieth, D., Gorska, A. E., Dumont, N., Shappell, S., Washington, M. K., Neilson, E. G., and Moses, H. L. (2004) Science 303 848-851 - PubMed
-
- Faler, B. J., Macsata, R. A., Plummer, D., Mishra, L., and Sidawy, A. N. (2006) Perspect. Vasc. Surg. Endovasc. Ther. 18 55-62 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
