

Review
doi: 10.1038/nri2327.
Immunomodulatory mast cells: negative, as well as positive, regulators of immunity
Affiliations
- PMID: 18483499
- PMCID: PMC2855166
- DOI: 10.1038/nri2327
Item in Clipboard
Review
Immunomodulatory mast cells: negative, as well as positive, regulators of immunity
Nat Rev Immunol.
2008 Jun.
Abstract
Mast cells can promote inflammation and other tissue changes in IgE-associated allergic disorders, as well as in certain innate and adaptive immune responses that are thought to be independent of IgE. However, mast cells can also have anti-inflammatory and immunosuppressive functions. Here, we review the evidence that mast cells can have negative, as well as positive, immunomodulatory roles in vivo, and we propose that mast cells can both enhance and later suppress certain features of an immune response.
Figures
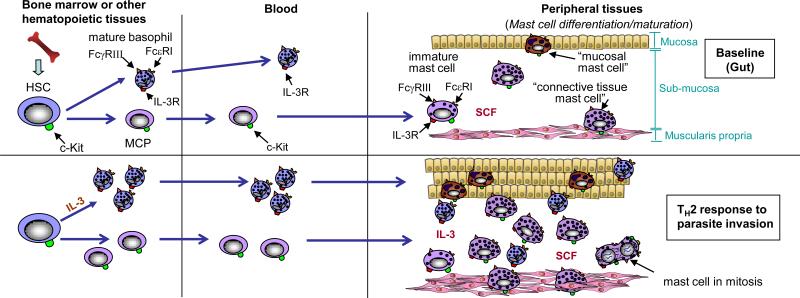
Tissue mast cells are derived from haematopoietic stem cells (HSCs), which ultimately give rise to mast-cell progenitors (MCPs). MCPs circulate in the blood and enter the tissues where they undergo differentiation and maturation to become mature mast cells. Stem-cell factor (SCF) ordinarily is required to maintain mast-cell survival, but the phenotype of mature mast cells can vary depending on the growth factor milieu (for example the presence or absence of additional cytokines with effects on mast-cell proliferation or phenotype, such as interleukin-3 (IL-3), IL-4, IL-9 and TGF-β1 and other microenvironmental factors. For example, mucosal mast cells are found in the mucosa of the gut, whereas connective tissue mast cells, that exhibit certain phenotypic characteristics that differ from those of mucosal mast cells, reside in the submucosa and muscularis propria. The numbers of these mast-cell populations can increase dramatically during a T helper 2 (TH2) type response to parasitic infection of the gut, which may reflect a combination of increased recruitment, survival and/or differentiation and maturation of MCPs, as well as proliferation of mast cells resident at that site.
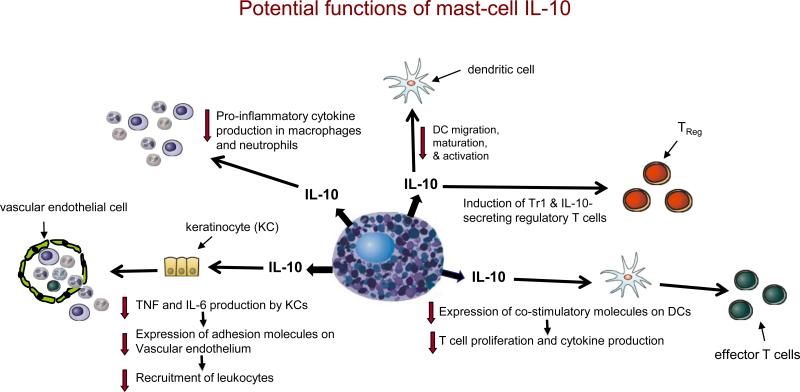
If produced in appropriate settings and amounts, mast-cell-derived interleukin-10 (IL-10) has the potential to promote the development of IL-10-secreting regulatory T cells (e.g., Tr1 cells and CD4+CD25+FOXP3+ T cells) and reduce DC migration, maturation and activation. It can also enhance the ability of DCs to reduce T-cell proliferation and cytokine production through the downregulation of costimulatory molecule expression by the DC. By directly inhibiting tumour-necrosis factor (TNF) and IL-6 production by keratinocytes, IL-10 can indirectly reduce the expression of adhesion molecules on vascular endothelial cells and thereby diminish the recruitment of circulating effector cells. IL-10 can directly inhibit the production of prostanoids by neutrophils and pro-inflammatory cytokines by macrophages. Although many of the specific functions indicated are based on evidence from in vitro studies of IL-10, mast-cell-derived IL-10 has been shown to mediate negative immunomodulatory functions in vivo.
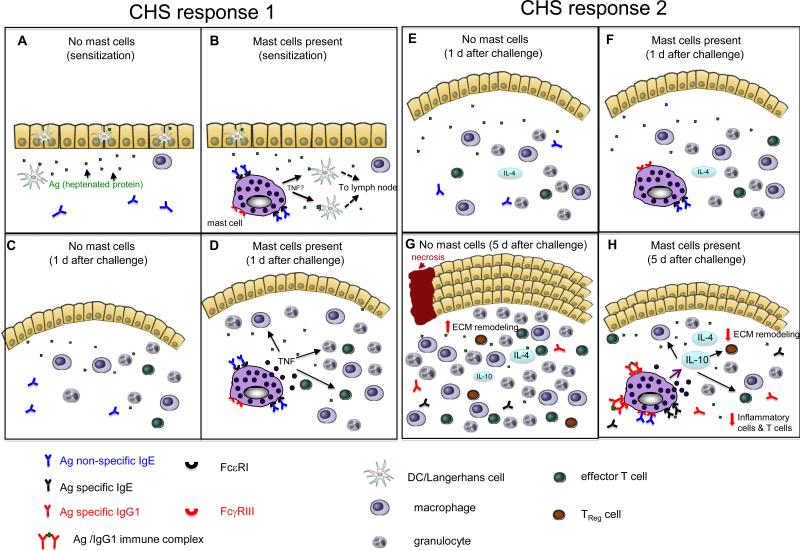
(a-d) CHS response 1 is elicited by sensitizing mice epicutaneously on the abdomen with 2% oxazolone in 100% ethanol and challenging them on the ear 5 d later with 1% oxazolone in 100% ethanol. In comparison to reactions elicited in the absence of mast cells (a, c), mast cells promote migration of dendritic cells (DCs) from the site of cutaneous sensitization (b) and, 1 day after epicutaneous challenge (that is day 6 after sensitization), mast cells promote features of the effector phase of the response, such as swelling of the dermis (indicated in pink in the figure) and leukocyte recruitment, by directly or indirectly undergoing antigen-dependent activation and releasing mediators (including tumour-necrosis factor (TNF)) (d). In CHS response 1, mast cells have a net pro-inflammatory effect, by enhancing both the sensitization and the effector phases of the response. The ability of mast cells to enhance sensitization requires that the mast cells have antigen non-specific IgE bound to their FcεRI. The mechanism of mast-cell activation during the effector phase of the response is not clear. (e-h) CHS response 2 is elicited by sensitizing mice epicutaneously on the abdomen with DNFB (0.5% vol/vol) in 100% acetone and challenging them on the ear 5 d later with DNFB (0.2% vol/vol) in 100% acetone. (e,g) One day after epicutaneous challenge with a hapten (that is day 6 after sensitization), the ear swelling responses are similar in the presence (f) or absence (e) of mast cells. Therefore, mast-cell activation probably does not have a crucial role early after challenge. However, five days after epicutaneous challenge with hapten (g,h), when the mice have elevated circulating levels of antigen-specific IgG1 antibodies, mast-cell-derived interleukin-10 (IL-10) contributes to the ability of mast cells to limit the number of innate inflammatory cells and T cells, and tissue pathology, at the site of hapten challenge (h). Based on in vitro studies, we speculate that increased local expression of certain cytokines (such as IL-4) at the site of hapten challenge can increase mast-cell surface expression of low-affinity Fc receptor for IgG (FcγRIII), and perhaps have other effects on mast-cell phenotype and/or function. Such mast cells can then secrete higher levels of TNF and IL-10 following stimulation through their FcγRIII by immune complexes of specific antigen and IgG1 antibodies. In the absence of mast cells (and mast-cell-derived IL-10), the pathology associated with these contact hypersensitivity responses is substantially exacerbated and there is increased inflammation (including higher numbers of CD8+, CD4+, and CD4+CD25+ T cells), more marked thickening of the epidermis, more substantial increases in extracellular matrix (ECM) and other components of the dermis, as well as areas of full thickness epidermal necrosis and ulceration (g). In CHS response 2, mast cells do not appear to enhance sensitization (not shown) but have a net anti-inflammatory effect on the effector phase of the response.
References
-
- Galli SJ, et al. Mast cells as “tunable” effector and immunoregulatory cells: recent advances. Annu Rev Immunol. 2005;23:749–786. - PubMed
-
- Mekori YA, Metcalfe DD. Mast cells in innate immunity. Immunol. Rev. 2000;173:131–140. - PubMed
-
- Dawicki W, Marshall JS. New and emerging roles for mast cells in host defence. Curr Opin Immunol. 2007;19:31–38. - PubMed
-
- Gilfillan AM, Tkaczyk C. Integrated signalling pathways for mast-cell activation. Nat Rev Immunol. 2006;6:218–230. - PubMed
-
- Grimbaldeston MA, Metz M, Yu M, Tsai M, Galli SJ. Effector and potential immunoregulatory roles of mast cells in IgE-associated acquired immune responses. Curr Opin Immunol. 2006;18:751–760. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
