

Granzyme A cleaves a mitochondrial complex I protein to initiate caspase-independent cell death
- PMID: 18485875
- PMCID: PMC2840390
- DOI: 10.1016/j.cell.2008.03.032
Granzyme A cleaves a mitochondrial complex I protein to initiate caspase-independent cell death
Abstract
The killer lymphocyte protease granzyme A (GzmA) triggers caspase-independent target cell death with morphological features of apoptosis. We previously showed that GzmA acts directly on mitochondria to generate reactive oxygen species (ROS) and disrupt the transmembrane potential (DeltaPsi(m)) but does not permeabilize the mitochondrial outer membrane. Mitochondrial damage is critical to GzmA-induced cell death since cells treated with superoxide scavengers are resistant to GzmA. Here we find that GzmA accesses the mitochondrial matrix to cleave the complex I protein NDUFS3, an iron-sulfur subunit of the NADH:ubiquinone oxidoreductase complex I, after Lys56 to interfere with NADH oxidation and generate superoxide anions. Target cells expressing a cleavage site mutant of NDUFS3 are resistant to GzmA-mediated cell death but remain sensitive to GzmB.
Figures
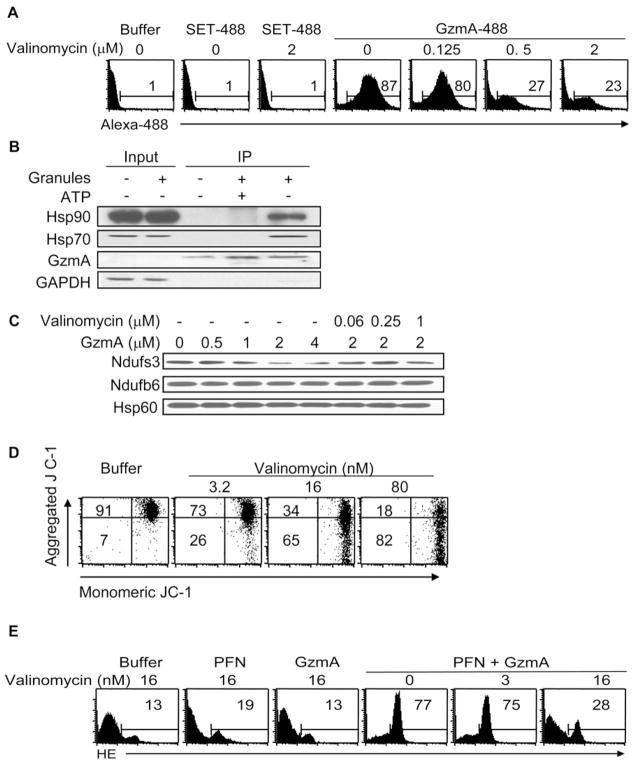
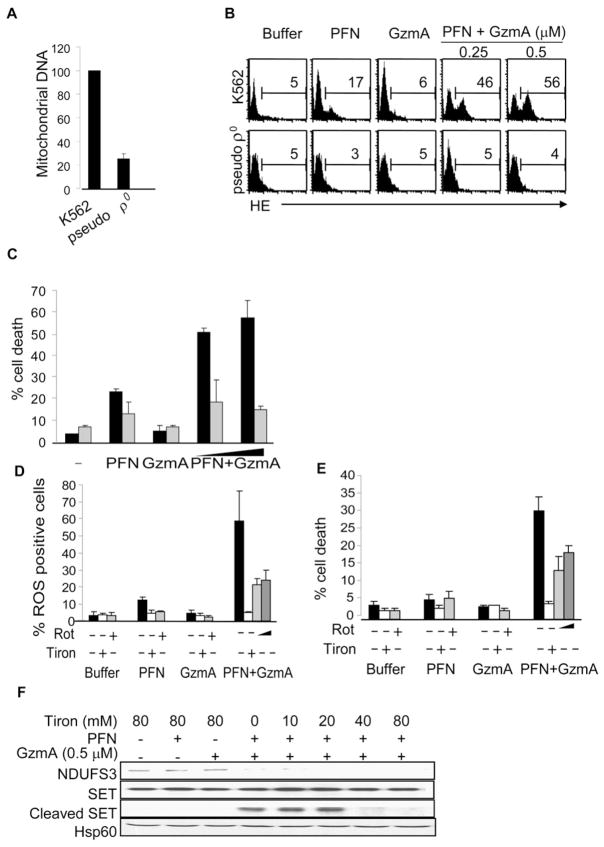
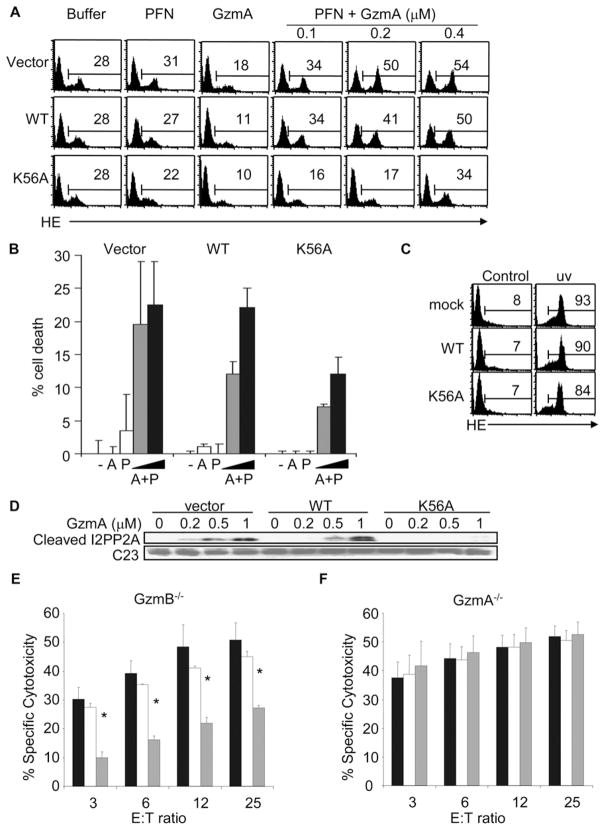
Comment in
-
Granzyme a, a stealth killer in the mitochondrion.Cell. 2008 May 16;133(4):568-70. doi: 10.1016/j.cell.2008.04.031. Cell. 2008. PMID: 18485862
Similar articles
-
Granzyme a, a stealth killer in the mitochondrion.Cell. 2008 May 16;133(4):568-70. doi: 10.1016/j.cell.2008.04.031. Cell. 2008. PMID: 18485862
-
Granzyme A activates another way to die.Immunol Rev. 2010 May;235(1):93-104. doi: 10.1111/j.0105-2896.2010.00902.x. Immunol Rev. 2010. PMID: 20536557 Free PMC article. Review.
-
Granzyme B-induced cell death exerted by ex vivo CTL: discriminating requirements for cell death and some of its signs.Cell Death Differ. 2008 Mar;15(3):567-79. doi: 10.1038/sj.cdd.4402289. Epub 2007 Dec 7. Cell Death Differ. 2008. PMID: 18064039
-
Apoptotic pathways are selectively activated by granzyme A and/or granzyme B in CTL-mediated target cell lysis.J Cell Biol. 2004 Nov 8;167(3):457-68. doi: 10.1083/jcb.200406115. J Cell Biol. 2004. PMID: 15534000 Free PMC article.
-
Granzymes and Mitochondria.Biochemistry (Mosc). 2020 Feb;85(2):131-139. doi: 10.1134/S0006297920020017. Biochemistry (Mosc). 2020. PMID: 32093590 Review.
Cited by
-
GZMA suppressed GPX4-mediated ferroptosis to improve intestinal mucosal barrier function in inflammatory bowel disease.Cell Commun Signal. 2024 Oct 4;22(1):474. doi: 10.1186/s12964-024-01836-y. Cell Commun Signal. 2024. PMID: 39367435 Free PMC article.
-
Combining Photodynamic Therapy and Targeted Drug Delivery Systems: Enhancing Mitochondrial Toxicity for Improved Cancer Outcomes.Int J Mol Sci. 2024 Oct 8;25(19):10796. doi: 10.3390/ijms251910796. Int J Mol Sci. 2024. PMID: 39409125 Free PMC article. Review.
-
Single-Fluorescent Protein Reporters Allow Parallel Quantification of Natural Killer Cell-Mediated Granzyme and Caspase Activities in Single Target Cells.Front Immunol. 2018 Aug 8;9:1840. doi: 10.3389/fimmu.2018.01840. eCollection 2018. Front Immunol. 2018. PMID: 30135688 Free PMC article.
-
The role of the mitochondria and the endoplasmic reticulum contact sites in the development of the immune responses.Cell Death Dis. 2018 Feb 28;9(3):336. doi: 10.1038/s41419-017-0237-7. Cell Death Dis. 2018. PMID: 29491398 Free PMC article. Review.
-
Granzyme A- and B-cluster deficiency delays acute lung injury in pneumovirus-infected mice.J Immunol. 2010 Jan 15;184(2):931-8. doi: 10.4049/jimmunol.0903029. Epub 2009 Dec 16. J Immunol. 2010. PMID: 20018616 Free PMC article.
References
-
- Aronis A, Melendez JA, Golan O, Shilo S, Dicter N, Tirosh O. Potentiation of Fas-mediated apoptosis by attenuated production of mitochondria-derived reactive oxygen species. Cell Death Differ. 2003;10:335–344. - PubMed
-
- Bell JK, Goetz DH, Mahrus S, Harris JL, Fletterick RJ, Craik CS. The oligomeric structure of human granzyme A is a determinant of its extended substrate specificity. Nat Struct Biol. 2003;10:527–534. - PubMed
-
- Beresford PJ, Jaju M, Friedman RS, Yoon MJ, Lieberman J. A role for heat shock protein 27 in CTL-mediated cell death. J Immunol. 1998;161:161–167. - PubMed
-
- Beresford PJ, Xia Z, Greenberg AH, Lieberman J. Granzyme A loading induces rapid cytolysis and a novel form of DNA damage independently of caspase activation. Immunity. 1999;10:585–594. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
