

Specificity, affinity and efficacy of iota-conotoxin RXIA, an agonist of voltage-gated sodium channels Na(V)1.2, 1.6 and 1.7
- PMID: 18486102
- PMCID: PMC2700742
- DOI: 10.1016/j.bcp.2008.03.019
Specificity, affinity and efficacy of iota-conotoxin RXIA, an agonist of voltage-gated sodium channels Na(V)1.2, 1.6 and 1.7
Abstract
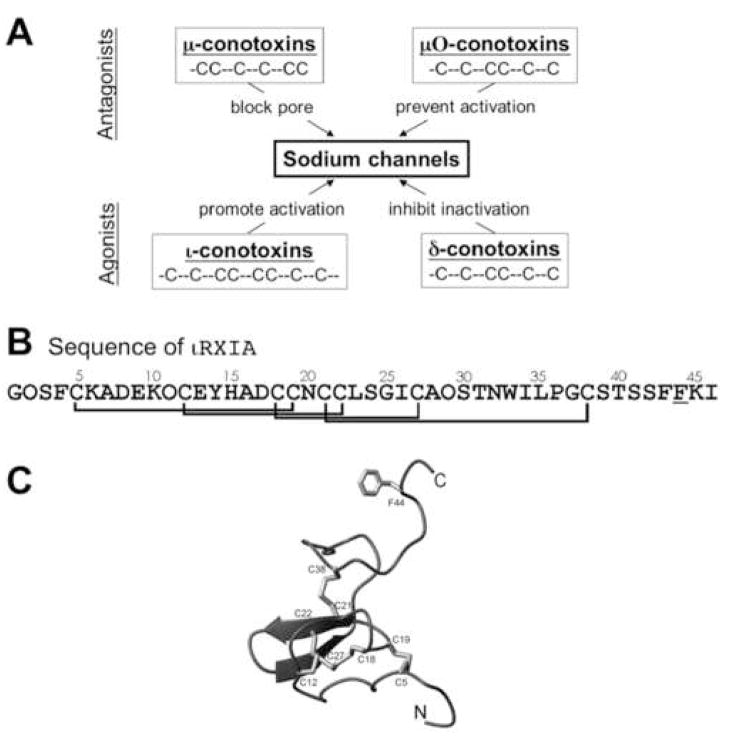
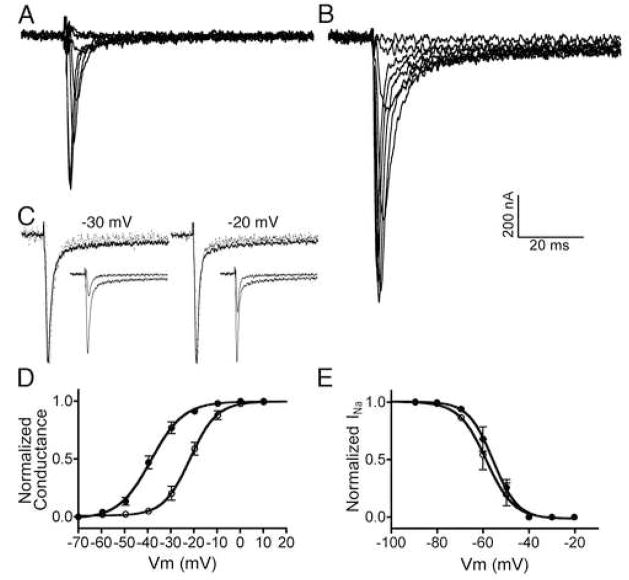
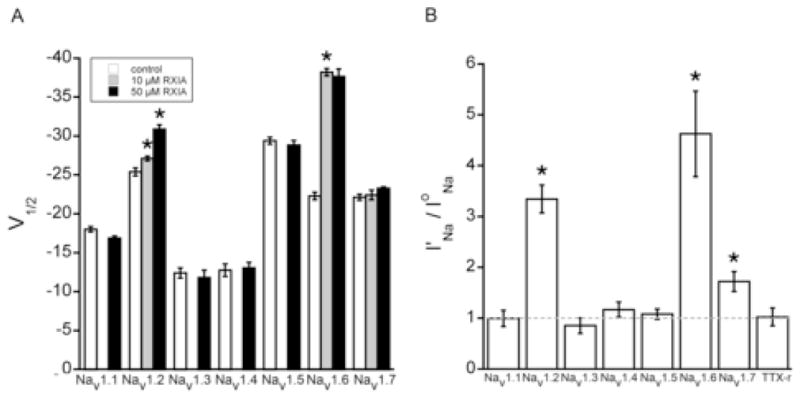
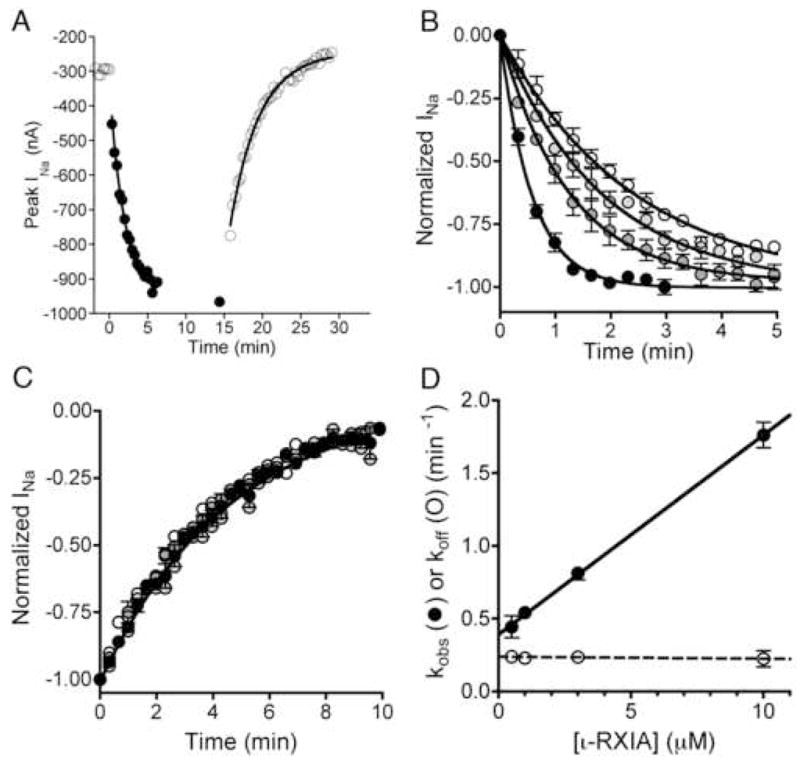
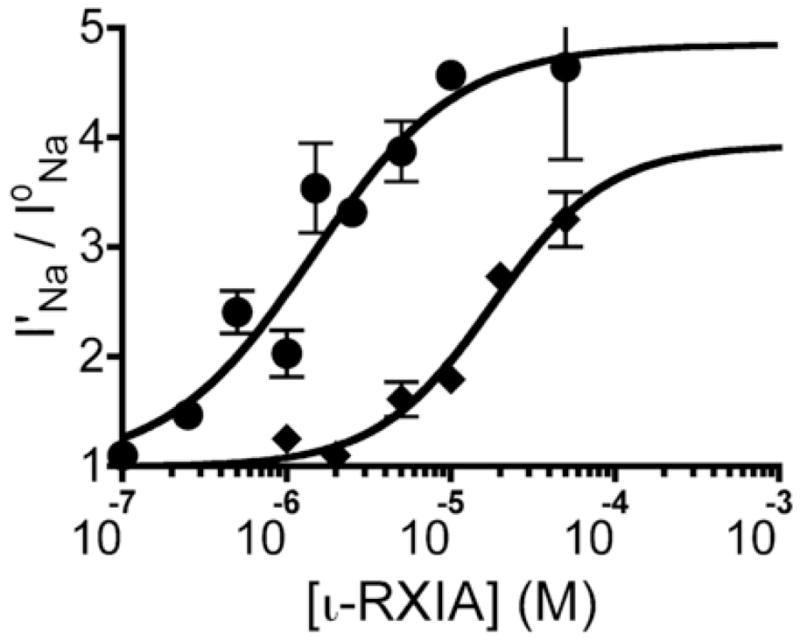
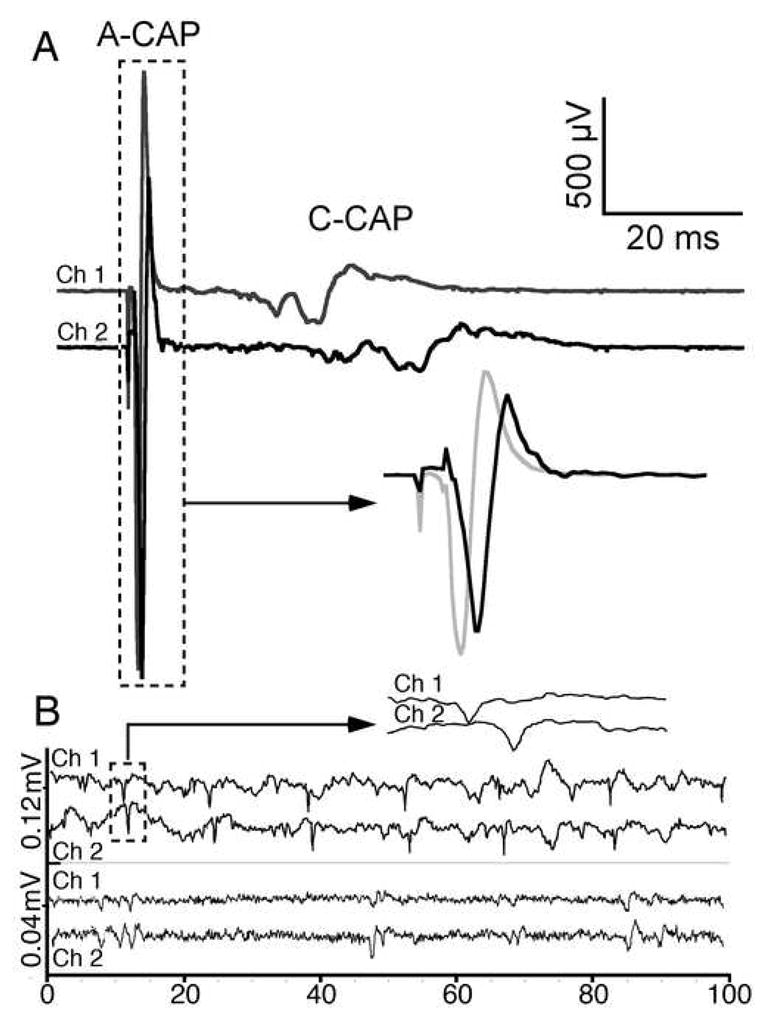
The excitotoxic conopeptide iota-RXIA induces repetitive action potentials in frog motor axons and seizures upon intracranial injection into mice. We recently discovered that iota-RXIA shifts the voltage-dependence of activation of voltage-gated sodium channel Na(V)1.6 to a more hyperpolarized level. Here, we performed voltage-clamp experiments to examine its activity against rodent Na(V)1.1 through Na(V)1.7 co-expressed with the beta1 subunit in Xenopus oocytes and Na(V)1.8 in dissociated mouse DRG neurons. The order of sensitivity to iota-RXIA was Na(V)1.6 > 1.2 > 1.7, and the remaining subtypes were insensitive. The time course of iota-RXIA-activity on Na(V)1.6 during exposure to different peptide concentrations were well fit by single-exponential curves that provided k(obs). The plot of k(obs)versus [iota-RXIA] was linear, consistent with a bimolecular reaction with a K(d) of approximately 3 microM, close to the steady-state EC(50) of approximately 2 microM. iota-RXIA has an unusual residue, D-Phe, and the analog with an L-Phe instead, iota-RXIA[L-Phe44], had a two-fold lower affinity and two-fold faster off-rate than iota-RXIA on Na(V)1.6 and furthermore was inactive on Na(V)1.2. iota-RXIA induced repetitive action potentials in mouse sciatic nerve with conduction velocities of both A- and C-fibers, consistent with the presence of Na(V)1.6 at nodes of Ranvier as well as in unmyelinated axons. Sixteen peptides homologous to iota-RXIA have been identified from a single species of Conus, so these peptides represent a rich family of novel sodium channel-targeting ligands.
Figures

References
-
- Gonzalez-Burgos G, Barrionuevo G. Voltage-gated sodium channels shape subthreshold EPSPs in layer 5 pyramidal neurons from rat prefrontal cortex. J Neurophysiol. 2001;86:1671–84. - PubMed
-
- Sontheimer H, Black JA, Waxman SG. Voltage-gated Na+ channels in glia: properties and possible functions. Trends Neurosci. 1996;19:325–31. - PubMed
-
- Catterall WA, Goldin AL, Waxman SG. International Union of Pharmacology. XLVII. Nomenclature and structure-function relationships of voltage-gated sodium channels. Pharmacol Rev. 2005;57:397–409. - PubMed
-
- Cestele S, Catterall WA. Molecular mechanisms of neurotoxin action on voltage-gated sodium channels. Biochimie. 2000;82:883–92. - PubMed
-
- Norton RS, Olivera BM. Conotoxins down under. Toxicon. 2006;48:780–98. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
