

Evolved physiological responses of phytoplankton to their integrated growth environment
- PMID: 18487129
- PMCID: PMC2606763
- DOI: 10.1098/rstb.2008.0019
Evolved physiological responses of phytoplankton to their integrated growth environment
Abstract
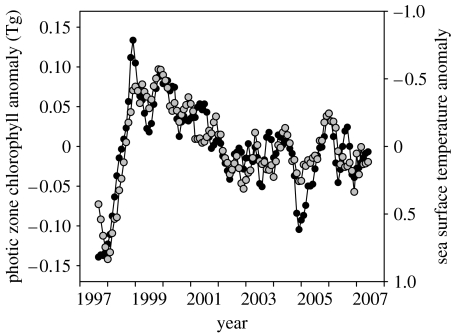
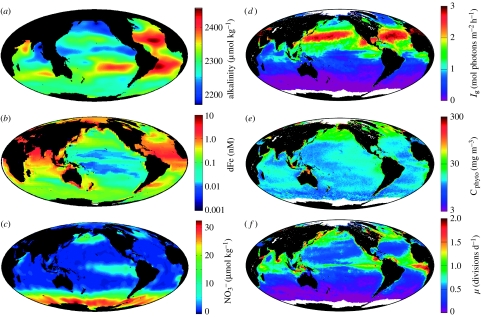
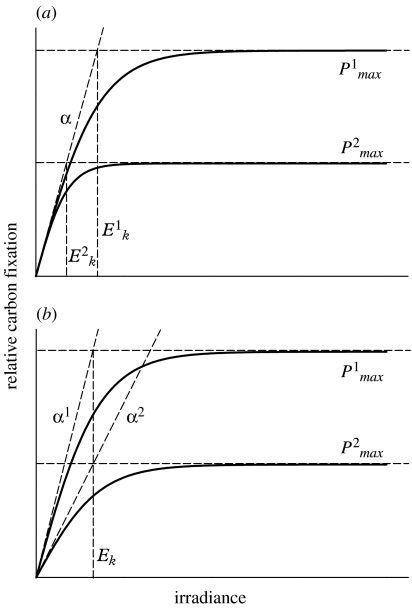
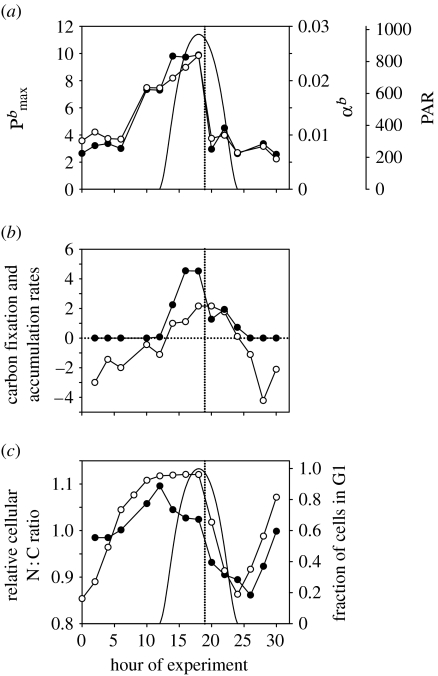
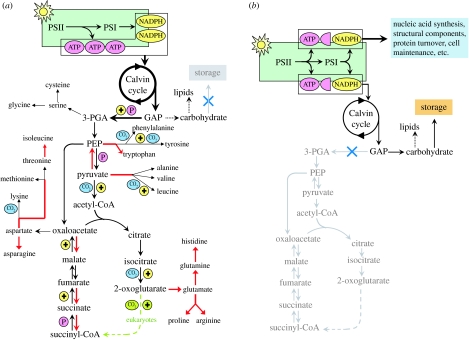
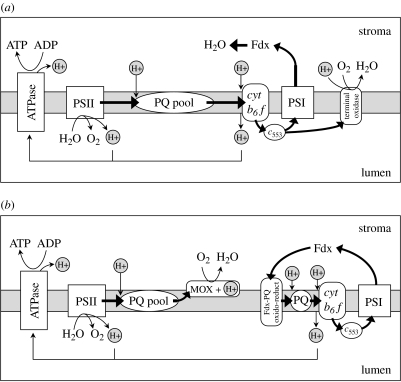
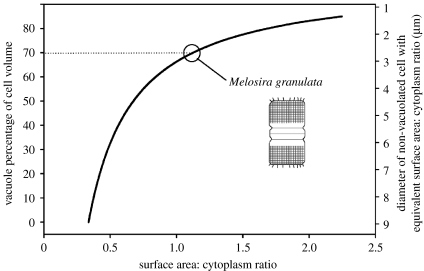
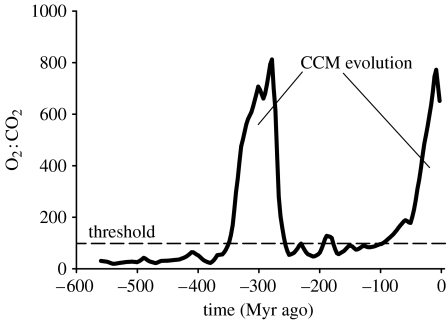
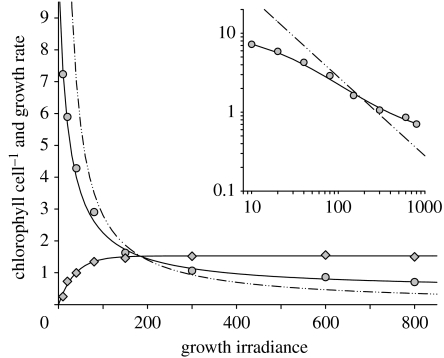
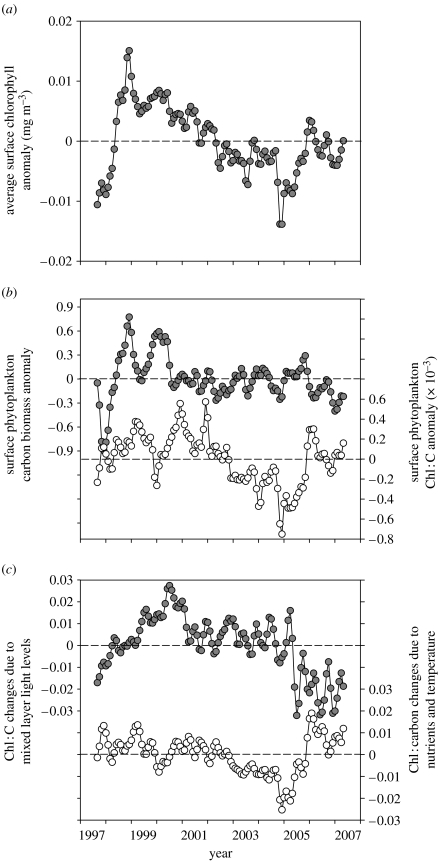
Phytoplankton growth and productivity relies on light, multiple nutrients and temperature. These combined factors constitute the 'integrated growth environment'. Since their emergence in the Archaean ocean, phytoplankton have experienced dramatic shifts in their integrated growth environment and, in response, evolved diverse mechanisms to maximize growth by optimizing the allocation of photosynthetic resources (ATP and NADPH) among all cellular processes. Consequently, co-limitation has become an omnipresent condition in the global ocean. Here we focus on evolved phytoplankton populations of the contemporary ocean and the varied energetic pathways they employ to solve the optimization problem of resource supply and demand. Central to this discussion is the allocation of reductant formed through photosynthesis, which we propose has the following three primary fates: carbon fixation, direct use and ATP generation. Investment of reductant among these three sinks is tied to cell cycle events, differentially influenced by specific forms of nutrient stress, and a strong determinant of relationships between light-harvesting (pigment), photosynthetic electron transport and carbon fixation. Global implications of optimization are illustrated by deconvolving trends in the 10-year global satellite chlorophyll record into contributions from biomass and physiology, thereby providing a unique perspective on the dynamic nature of surface phytoplankton populations and their link to climate.
Figures
References
-
- Asada K. The water–water cycle in chloroplasts: scavenging of active oxygens and dissipation of excess photons. Annu. Rev. Plant Physiol. Plant Mol. Biol. 1999;50:601–639. doi:10.1146/annurev.arplant.50.1.601 - DOI - PubMed
-
- Badger M.R, Price G.D. CO2 concentrating mechanisms in cyanobacteria: molecular components, their diversity and evolution. J. Exp. Bot. 2003;54:609–622. doi:10.1093/jxb/erg076 - DOI - PubMed
-
- Badger M.R, Andrews T.J, Whitney S.M, Ludwig M, Yellowlees D.C, Leggat W, Price G.D. The diversity and coevolution of rubisco, plastids, pyrenoids, and chloroplast-based CO2-concentrating mechanisms in algae. Can. J. Bot. 1998;76:1052–1071. doi:10.1139/cjb-76-6-1052 - DOI
-
- Bailey, S. et al In press. Photosynthesis in marine Synechococcus and the critical nature of electron flow to oxygen. Biochim. Biophys. Acta - PubMed
-
- Behrenfeld M.J, Falkowski P.G. Photosynthetic rates derived from satellite-based chlorophyll concentration. Limnol. Oceanogr. 1997;42:1–20.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources