

Molecular dynamics simulation of Kv channel voltage sensor helix in a lipid membrane with applied electric field
- PMID: 18487312
- PMCID: PMC2483744
- DOI: 10.1529/biophysj.108.130658
Molecular dynamics simulation of Kv channel voltage sensor helix in a lipid membrane with applied electric field
Abstract
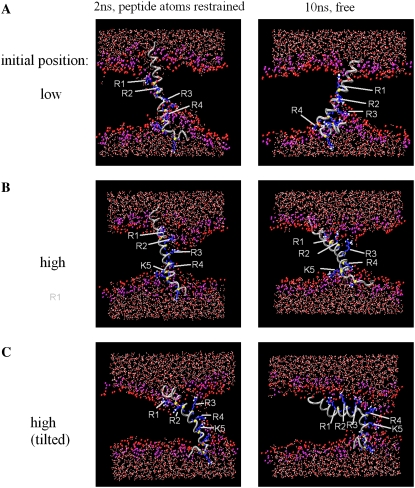
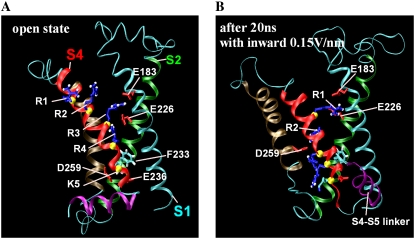
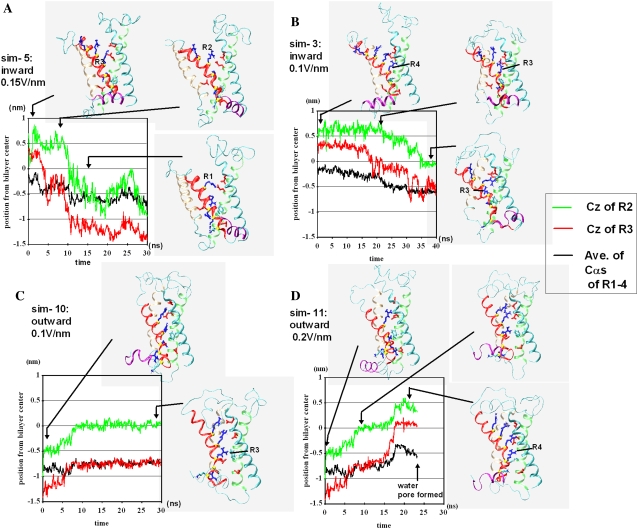
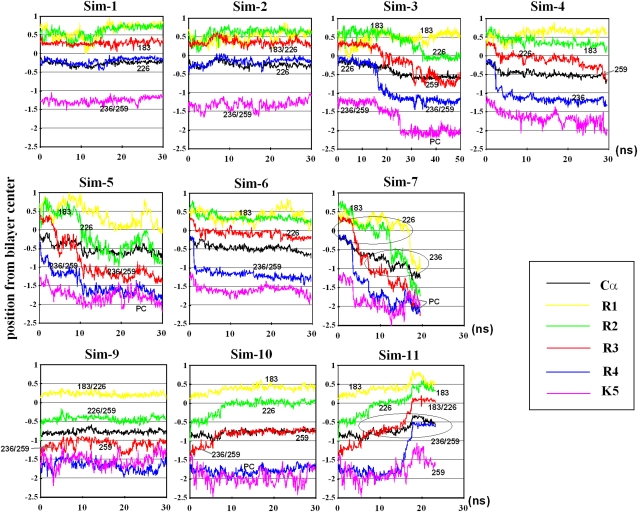
In this article, we present the results of the molecular dynamics simulations of amphiphilic helix peptides of 13 amino-acid residues, placed at the lipid-water interface of dipalmitoylphosphatidylcholine bilayers. The peptides are identical with, or are derivatives of, the N-terminal segment of the S4 helix of voltage-dependent K channel KvAP, containing four voltage-sensing arginine residues (R1-R4). Upon changing the direction of the externally applied electric field, the tilt angle of the wild-type peptide changes relative to the lipid-water interface, with the N-terminus heading up with an outward electric field. These movements were not observed using an octane membrane in place of the dipalmitoylphosphatidylcholine membrane, and were markedly suppressed by 1), substituting Phe located one residue before the first arginine (R1) with a hydrophilic residue (Ser, Thr); or 2), changing the periodicity rule of Rs from at-every-third to at-every-fourth position; or 3), replacing R1 with a lysine residue (K). These and other findings suggest that the voltage-dependent movement requires deep positioning of Rs when the resting (inward) electric field is present. Later, we performed simulations of the voltage sensor domain (S1-S4) of Kv1.2 channel. In simulations with a strong electric field (0.1 V/nm or above) and positional restraints on the S1 and S2 helices, S4 movement was observed consisting of displacement along the S4 helix axis and a screwlike axial rotation. Gating-charge-carrying Rs were observed to make serial interactions with E183 in S1 and E226 in S2, in the outer water crevice. A 30-ns-backward simulation started from the open-state model gave rise to a structure similar to the recent resting-state model, with S4 moving vertically approximately 6.7 A. The energy landscape around the movement of S4 appears very ragged due to salt bridges formed between gating-charge-carrying residues and negatively charged residues of S1, S2, and S3 helices. Overall, features of S3 and S4 movements are consistent with the recent helical-screw model. Both forward and backward simulations show the presence of at least two stable intermediate structures in which R2 and R3 form salt bridges with E183 or E226, respectively. These structures are the candidates for the states postulated in previous gating kinetic models, such as the Zagotta-Hoshi-Aldrich model, to account for more than one transition step per subunit for activation.
Figures


References
-
- Hille, B. 2001. Ion Channels of Excitable Membranes, 3rd Ed. Sinauer, Sunderland, MA.
-
- Jiang, Y., A. Lee, J. Chen, V. Ruta, M. Cadene, B. T. Chait, and R. MacKinnon. 2003. X-ray structure of a voltage-dependent K+ channel. Nature. 423:33–41. - PubMed
-
- Long, S. B., E. B. Campbell, and R. Mackinnon. 2005. Crystal structure of a mammalian voltage-dependent Shaker family K+ channel. Science. 309:897–903. - PubMed
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous
