

Mutation of a rice gene encoding a phenylalanine biosynthetic enzyme results in accumulation of phenylalanine and tryptophan
- PMID: 18487352
- PMCID: PMC2438470
- DOI: 10.1105/tpc.107.057455
Mutation of a rice gene encoding a phenylalanine biosynthetic enzyme results in accumulation of phenylalanine and tryptophan
Abstract
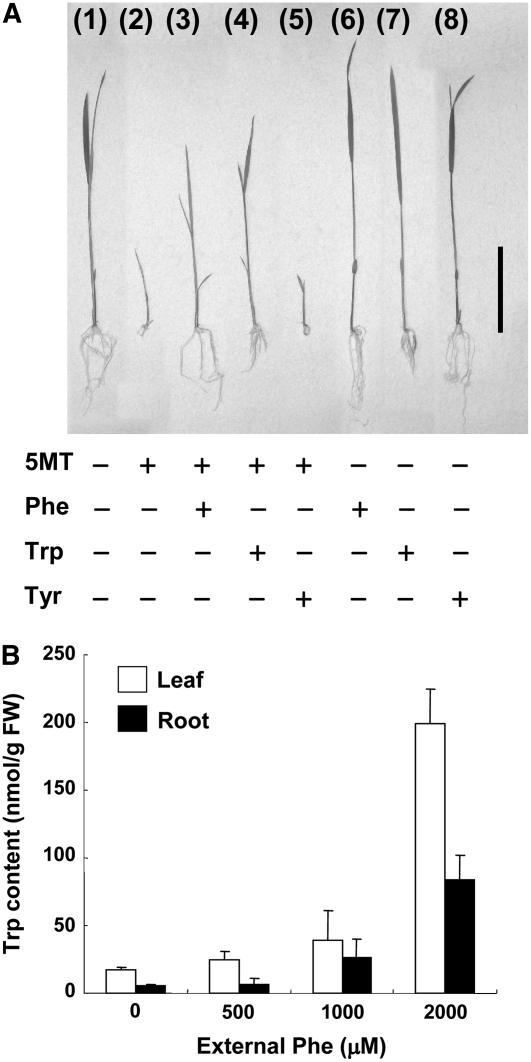
Two distinct biosynthetic pathways for Phe in plants have been proposed: conversion of prephenate to Phe via phenylpyruvate or arogenate. The reactions catalyzed by prephenate dehydratase (PDT) and arogenate dehydratase (ADT) contribute to these respective pathways. The Mtr1 mutant of rice (Oryza sativa) manifests accumulation of Phe, Trp, and several phenylpropanoids, suggesting a link between the synthesis of Phe and Trp. Here, we show that the Mtr1 mutant gene (mtr1-D) encodes a form of rice PDT with a point mutation in the putative allosteric regulatory region of the protein. Transformed callus lines expressing mtr1-D exhibited all the characteristics of Mtr1 callus tissue. Biochemical analysis revealed that rice PDT possesses both PDT and ADT activities, with a preference for arogenate as substrate, suggesting that it functions primarily as an ADT. The wild-type enzyme is feedback regulated by Phe, whereas the mutant enzyme showed a reduced feedback sensitivity, resulting in Phe accumulation. In addition, these observations indicate that rice PDT is critical for regulating the size of the Phe pool in plant cells. Feeding external Phe to wild-type callus tissue and seedlings resulted in Trp accumulation, demonstrating a connection between Phe accumulation and Trp pool size.
Figures









References
-
- Bate, N.J., Orr, J., Ni, W., Meromi, A., Nadler-Hassar, T., Doerner, P.W., Dixon, R.A., Lamb, C.J., and Elkind, Y. (1994). Quantitative relationship between phenylalanine ammonia-lyase levels and phenylpropanoid accumulation in transgenic tobacco identifies a rate-determining step in natural product synthesis. Proc. Natl. Acad. Sci. USA 91 7608–7612. - PMC - PubMed
-
- Bongaerts, J., Kramer, M., Muller, U., Raeven, L., and Wubbolts, M. (2001). Metabolic engineering for microbial production of aromatic amino acids and derived compounds. Metab. Eng. 3 289–300. - PubMed
-
- Bonner, C., and Jensen, R. (1987). Prephenate aminotransferase. Methods Enzymol. 142 479–487. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
