

Cyclic-di-GMP regulates extracellular polysaccharide production, biofilm formation, and rugose colony development by Vibrio vulnificus
- PMID: 18487410
- PMCID: PMC2446529
- DOI: 10.1128/AEM.00176-08
Cyclic-di-GMP regulates extracellular polysaccharide production, biofilm formation, and rugose colony development by Vibrio vulnificus
Abstract
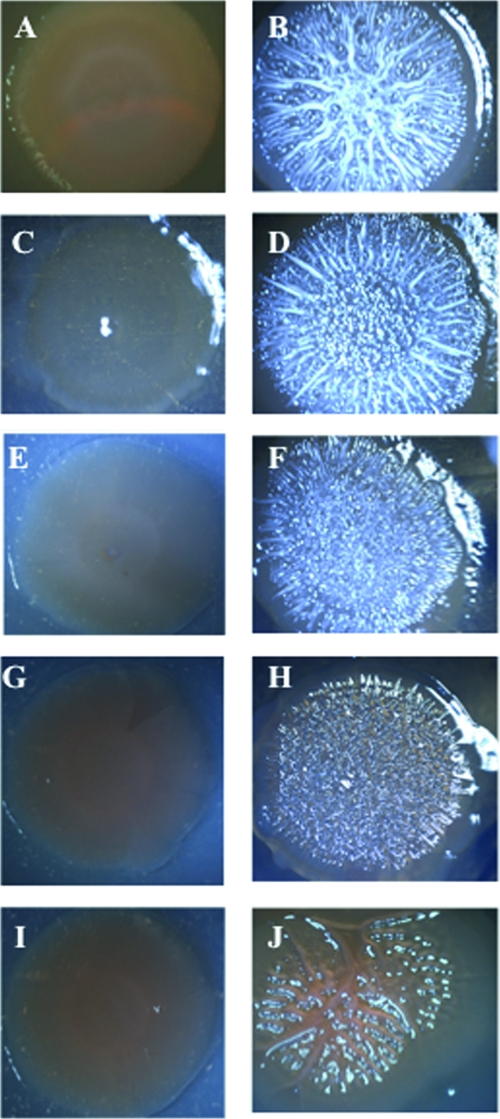
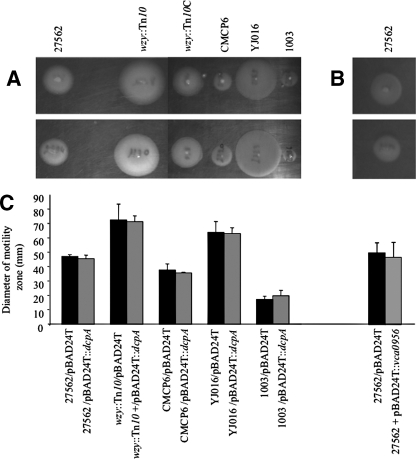
Vibrio vulnificus is a human and animal pathogen that carries the highest death rate of any food-borne disease agent. It colonizes shellfish and forms biofilms on the surfaces of plankton, algae, fish, and eels. Greater understanding of biofilm formation by the organism could provide insight into approaches to decrease its load in filter feeders and on biotic surfaces and control the occurrence of invasive disease. The capsular polysaccharide (CPS), although essential for virulence, is not required for biofilm formation under the conditions used here. In other bacteria, increased biofilm formation often correlates with increased exopolysaccharide (EPS) production. We exploited the translucent phenotype of acapsular mutants to screen a V. vulnificus genomic library and identify genes that imparted an opaque phenotype to both CPS biosynthesis and transport mutants. One of these encoded a diguanylate cyclase (DGC), an enzyme that synthesizes bis-(3'-5')-cyclic-di-GMP (c-di-GMP). This prompted us to use this DGC, DcpA, to examine the effect of elevated c-di-GMP levels on several developmental pathways in V. vulnificus. Increased c-di-GMP levels induced the production of an EPS that was distinct from the CPS and dramatically enhanced biofilm formation and rugosity in a CPS-independent manner. However, the EPS could not compensate for the loss of CPS production that is required for virulence. In contrast to V. cholerae, motility and virulence appeared unaffected by elevated levels of c-di-GMP.
Figures


References
-
- Bhasin, M., A. Garg, and G. P. Raghava. 2005. PSLpred: prediction of subcellular localization of bacterial proteins. Bioinformatics 21:2522-2524. - PubMed
-
- Bisharat, N., V. Agmon, R. Finkelstein, R. Raz, G. Ben-Dror, L. Lerner, S. Soboh, R. Colodner, D. N. Cameron, D. L. Wykstra, D. L. Swerdlow, and J. J. Farmer III. 1999. Clinical, epidemiological, and microbiological features of Vibrio vulnificus biogroup 3 causing outbreaks of wound infection and bacteraemia in Israel. Lancet 354:1421-1424. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
