

Consequences of lipid droplet coat protein downregulation in liver cells: abnormal lipid droplet metabolism and induction of insulin resistance
- PMID: 18487449
- PMCID: PMC2494696
- DOI: 10.2337/db07-1383
Consequences of lipid droplet coat protein downregulation in liver cells: abnormal lipid droplet metabolism and induction of insulin resistance
Abstract
Objective: Accumulation of intracellular lipid droplets (LDs) in non-adipose tissues is recognized as a strong prognostic factor for the development of insulin resistance in obesity. LDs are coated with perilipin, adipose differentiation-related protein, tail interacting protein of 47 kd (PAT) proteins that are thought to regulate LD turnover by modulating lipolysis. Our hypothesis is that PAT proteins modulate LD metabolism and therefore insulin resistance.
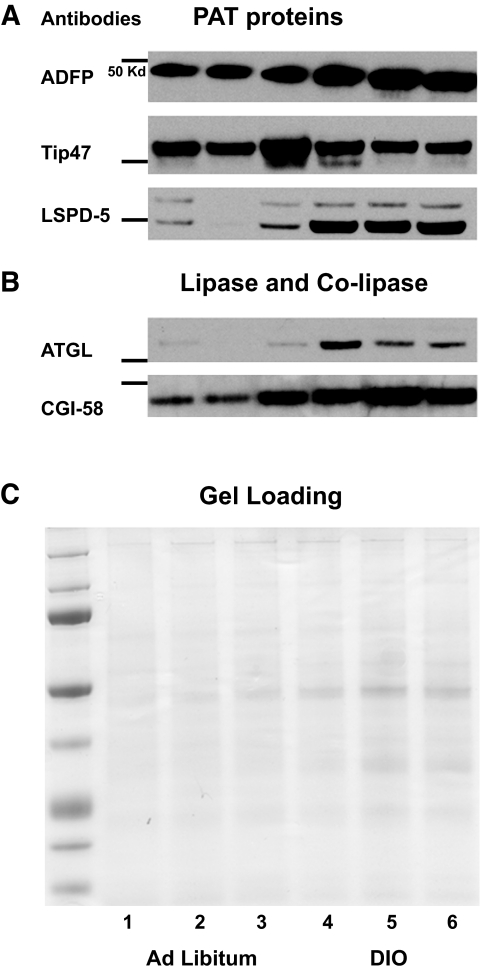
Research design and methods: We used a cell culture model (murine AML12 loaded with oleic acid) and small interfering RNA to directly assess the impact of PAT proteins on LD accumulation, lipid metabolism, and insulin action. PAT proteins associated with excess fat deposited in livers of diet-induced obese (DIO) mice were also measured.
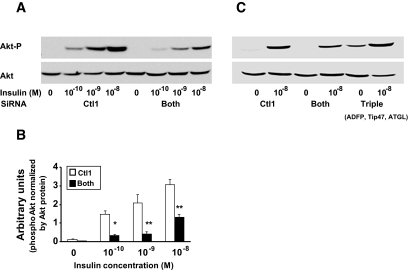
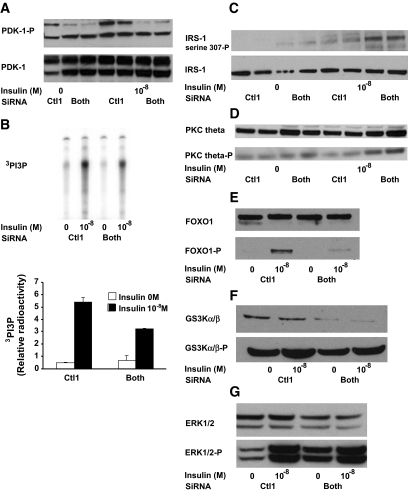
Results: Cells lacking PAT proteins exhibited a dramatic increase in LD size and a decrease in LD number. Further, the lipolytic rate increased by approximately 2- to 2.5-fold in association with increased adipose triglyceride lipase (ATGL) at the LD surface. Downregulation of PAT proteins also produced insulin resistance, as indicated by decreased insulin stimulation of Akt phosphorylation (P < 0.001). Phosphoinositide-dependent kinase-1 and phosphoinositide 3-kinase decreased, and insulin receptor substrate-1 307 phosphorylation increased. Increased lipids in DIO mice livers were accompanied by changes in PAT composition but also increased ATGL, suggesting a relative PAT deficiency.
Conclusions: These data establish an important role for PAT proteins as surfactant at the LD surface, packaging lipids in smaller units and restricting access of lipases and thus preventing insulin resistance. We suggest that a deficiency of PAT proteins relative to the quantity of ectopic fat could contribute to cellular dysfunction in obesity and type 2 diabetes.
Figures
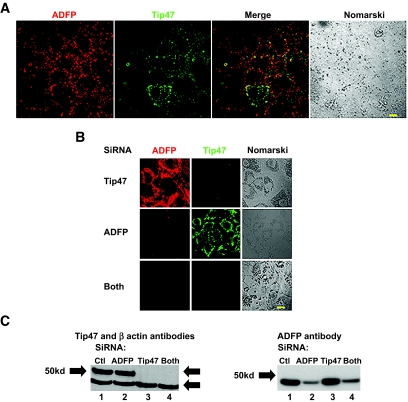
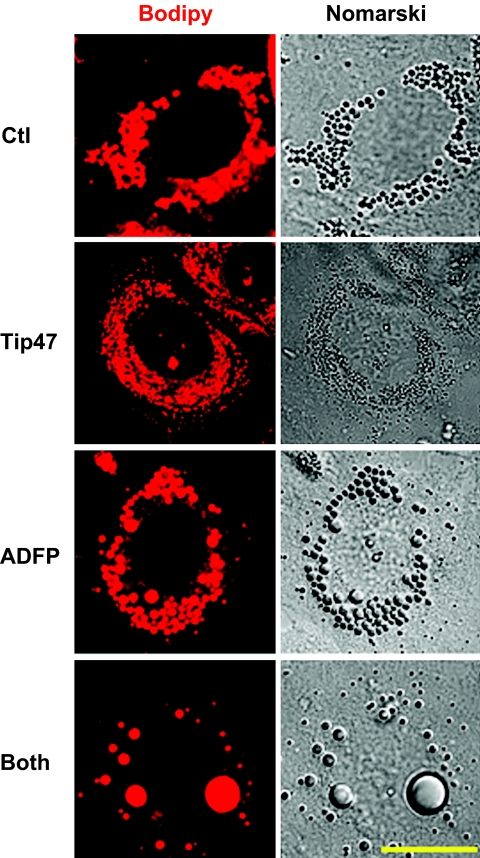
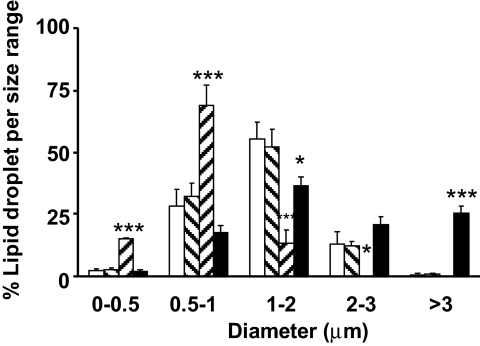
 ). Data are means ± SE from 11 separate experiments for control and ADFP, from three separate experiments for Tip47, and from 27 separate experiments for both. ***P < 0.0001; *P < 0.05 vs. control (t test).
). Data are means ± SE from 11 separate experiments for control and ADFP, from three separate experiments for Tip47, and from 27 separate experiments for both. ***P < 0.0001; *P < 0.05 vs. control (t test).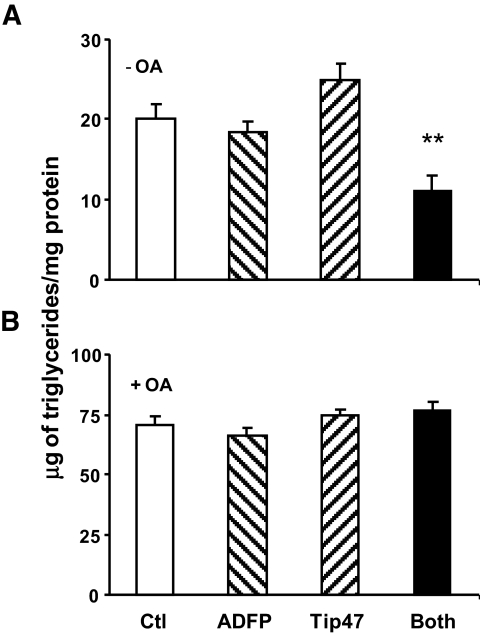 ). B: Triglycerides were measured in cells grown in culture media supplemented overnight with 400 μmol/l of oleic acid in cells treated as described above. Data are means ± SE from three separate experiments and P < 0.001 (one-way ANOVA) for cells lacking both ADFP and Tip47 grown without exogenous NEFA addition.
). B: Triglycerides were measured in cells grown in culture media supplemented overnight with 400 μmol/l of oleic acid in cells treated as described above. Data are means ± SE from three separate experiments and P < 0.001 (one-way ANOVA) for cells lacking both ADFP and Tip47 grown without exogenous NEFA addition.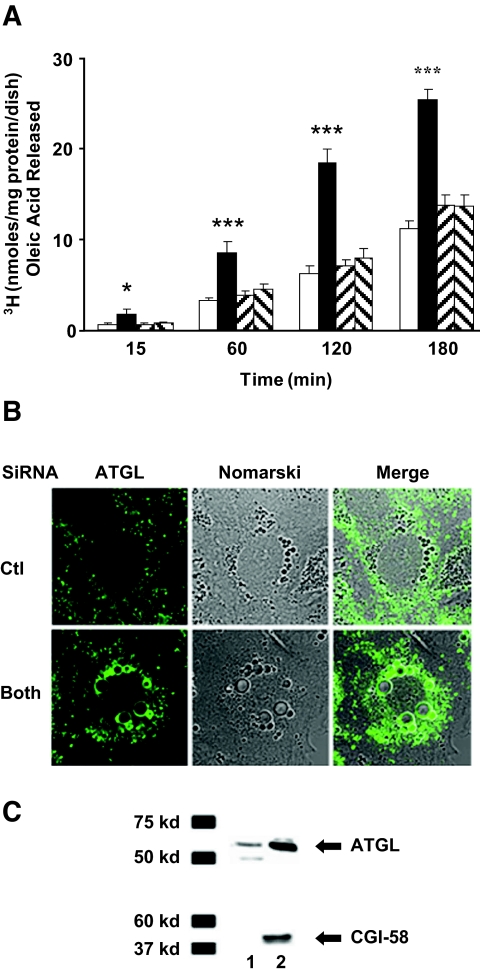
References
-
- Meigs JB: Epidemiology of the metabolic syndrome. Am J Manag Care 8 (Suppl. 11): S283–S292, 2002 - PubMed
-
- Eckel RH, Grundy SM, Zimmet PZ: The metabolic syndrome. Lancet 365: 1415–428, 2005 - PubMed
-
- Gavrilova O, Haluzik M, Matsusue K, Cutson JJ, Johnson L, Dietz KR, Nicole CJ, Vinson C, Gonzalez FJ, Reitman ML: Liver peroxisome proliferator-activated receptor gamma contributes to hepatic steatosis, triglyceride clearance, and regulation of body fat mass. J Biol Chem 278: 34268–34276, 2003 - PubMed
-
- Prentki M, Vischer S, Glennon MC, Regazzi R, Deeney JT, Corkey BE: Malonyl-CoA and long chain acyl-CoA esters as metabolic coupling factors in nutrient-induced insulin secretion. J Biol Chem 267: 5802–5810, 1992 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
