

STIM1 signalling controls store-operated calcium entry required for development and contractile function in skeletal muscle
- PMID: 18488020
- PMCID: PMC2694045
- DOI: 10.1038/ncb1731
STIM1 signalling controls store-operated calcium entry required for development and contractile function in skeletal muscle
Abstract
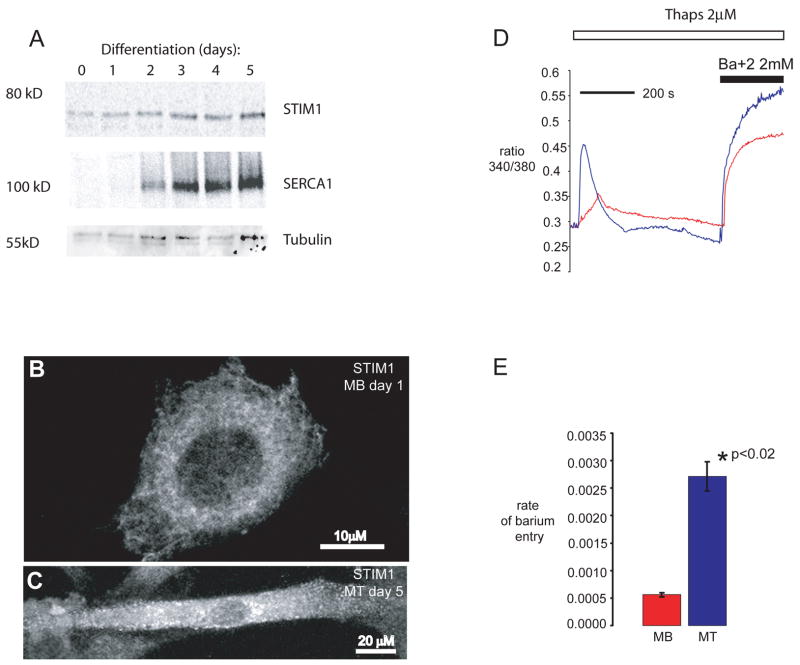
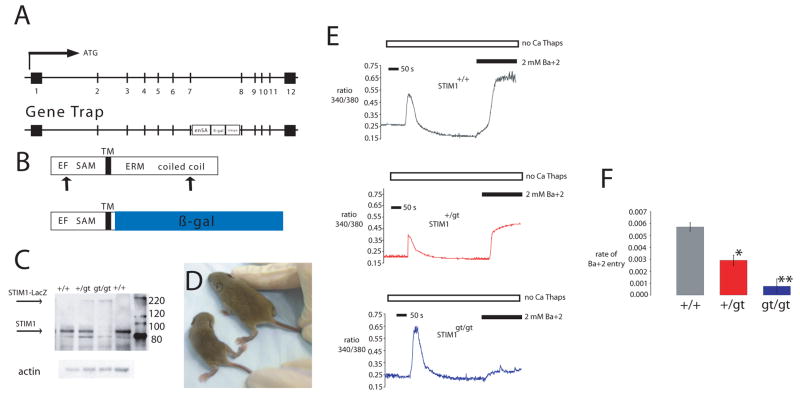
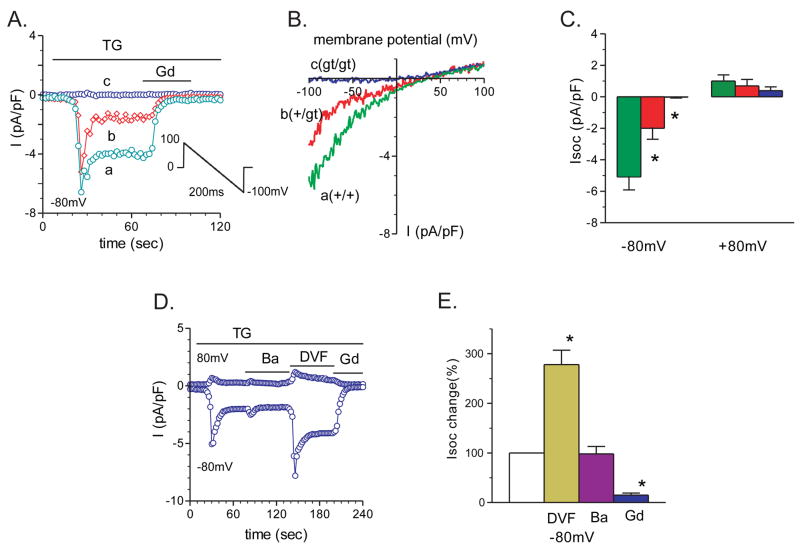
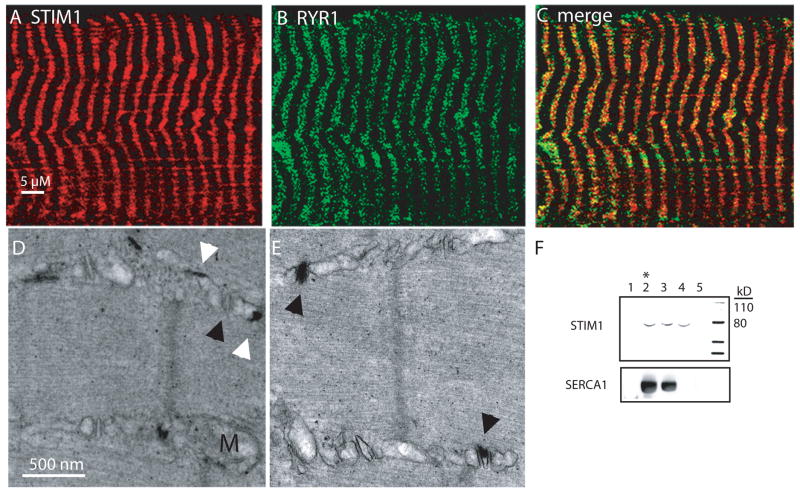
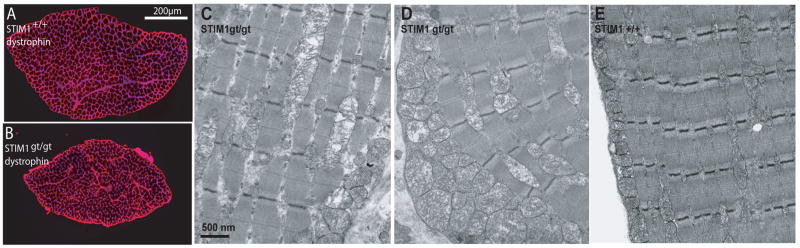
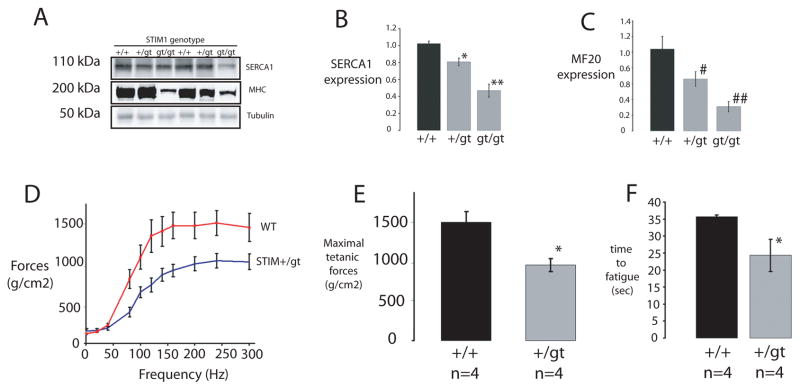
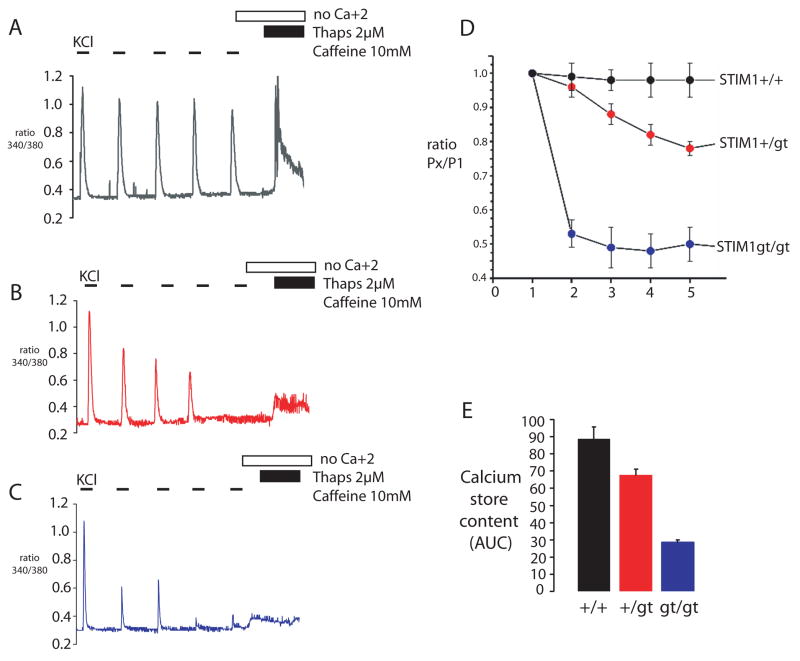
It is now well established that stromal interaction molecule 1 (STIM1) is the calcium sensor of endoplasmic reticulum stores required to activate store-operated calcium entry (SOC) channels at the surface of non-excitable cells. However, little is known about STIM1 in excitable cells, such as striated muscle, where the complement of calcium regulatory molecules is rather disparate from that of non-excitable cells. Here, we show that STIM1 is expressed in both myotubes and adult skeletal muscle. Myotubes lacking functional STIM1 fail to show SOC and fatigue rapidly. Moreover, mice lacking functional STIM1 die perinatally from a skeletal myopathy. In addition, STIM1 haploinsufficiency confers a contractile defect only under conditions where rapid refilling of stores would be needed. These findings provide insight into the role of STIM1 in skeletal muscle and suggest that STIM1 has a universal role as an ER/SR calcium sensor in both excitable and non-excitable cells.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
