

Opalin, a transmembrane sialylglycoprotein located in the central nervous system myelin paranodal loop membrane
- PMID: 18490449
- PMCID: PMC3258930
- DOI: 10.1074/jbc.M801314200
Opalin, a transmembrane sialylglycoprotein located in the central nervous system myelin paranodal loop membrane
Abstract
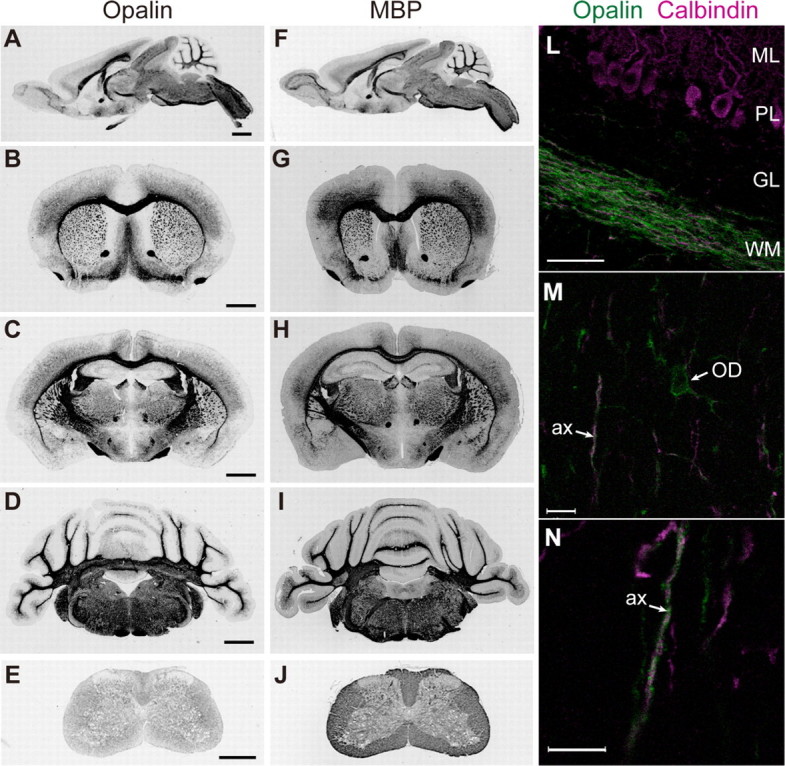
In contrast to compact myelin, the series of paranodal loops located in the outermost lateral region of myelin is non-compact; the intracellular space is filled by a continuous channel of cytoplasm, the extracellular surfaces between neighboring loops keep a definite distance, but the loop membranes have junctional specializations. Although the proteins that form compact myelin have been well studied, the protein components of paranodal loop membranes are not fully understood. This report describes the biochemical characterization and expression of Opalin as a novel membrane protein in paranodal loops. Mouse Opalin is composed of a short N-terminal extracellular domain (amino acid residues 1-30), a transmembrane domain (residues 31-53), and a long C-terminal intracellular domain (residues 54-143). Opalin is enriched in myelin of the central nervous system, but not that of the peripheral nervous system of mice. Enzymatic deglycosylation showed that myelin Opalin contained N- and O-glycans, and that the O-glycans, at least, had negatively charged sialic acids. We identified two N-glycan sites at Asn-6 and Asn-12 and an O-glycan site at Thr-14 in the extracellular domain. Site-directed mutations at the glycan sites impaired the cell surface localization of Opalin. In addition to the somata and processes of oligodendrocytes, Opalin immunoreactivity was observed in myelinated axons in a spiral fashion, and was concentrated in the paranodal loop region. Immunogold electron microscopy demonstrated that Opalin was localized at particular sites in the paranodal loop membrane. These results suggest a role for highly sialylglycosylated Opalin in an intermembranous function of the myelin paranodal loops in the central nervous system.
Figures






References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
