

Differential regulation of interleukin 12 and interleukin 23 production in human dendritic cells
- PMID: 18490488
- PMCID: PMC2413040
- DOI: 10.1084/jem.20071450
Differential regulation of interleukin 12 and interleukin 23 production in human dendritic cells
Abstract
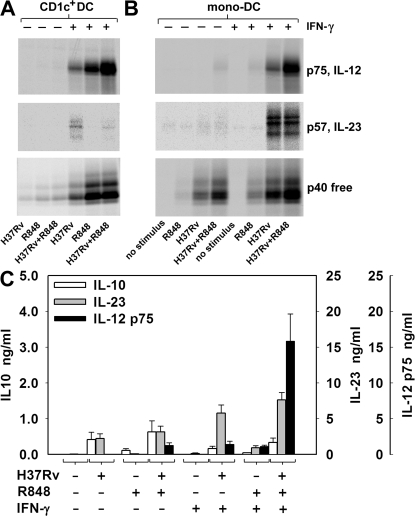
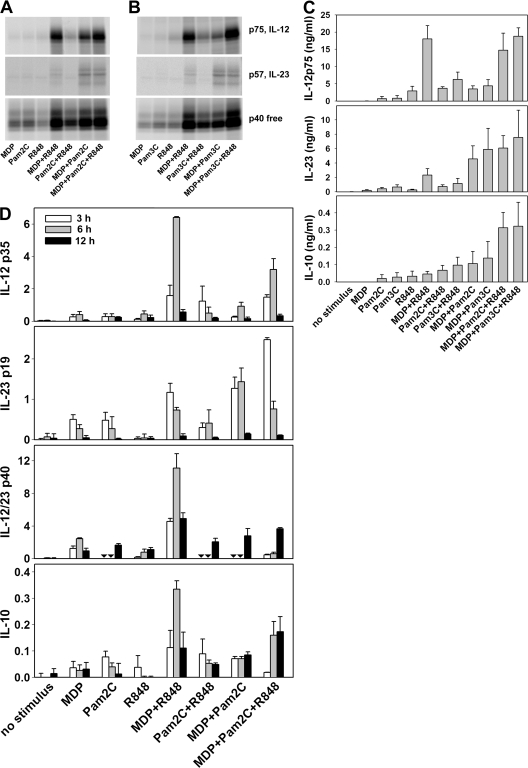
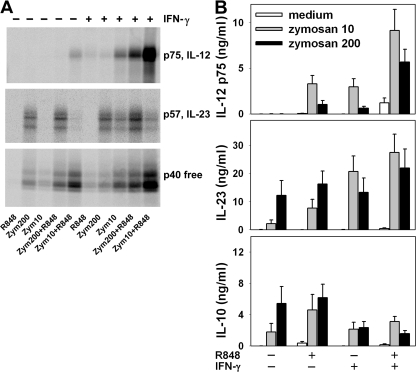
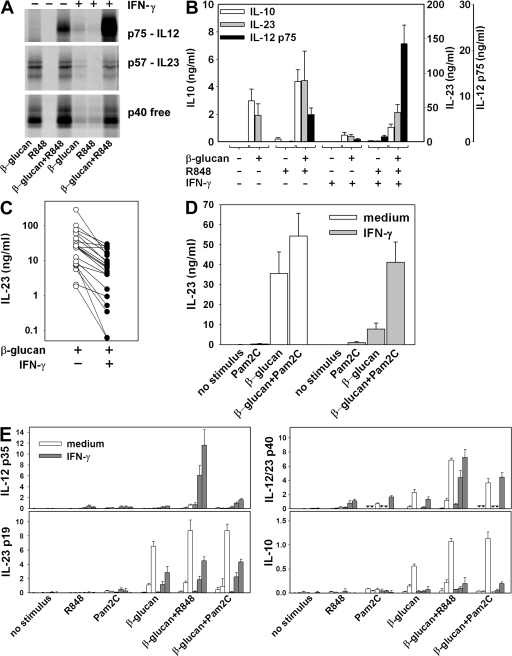
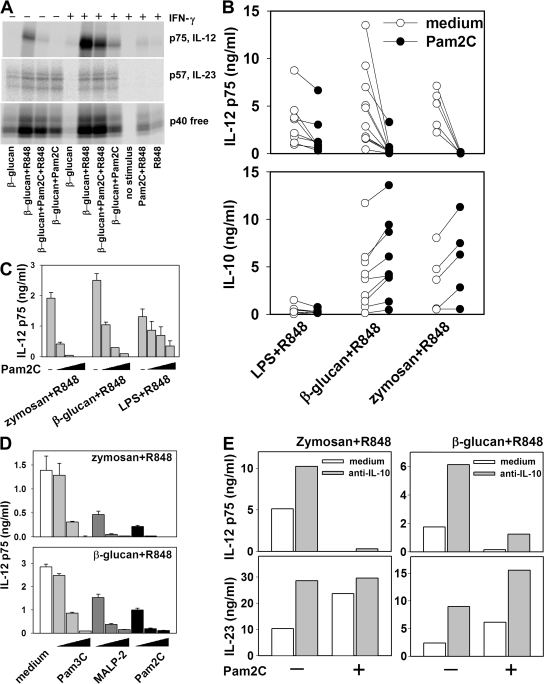
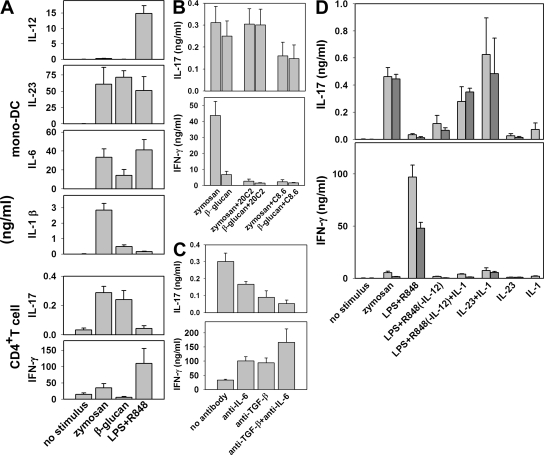
We analyzed interleukin (IL) 12 and IL-23 production by monocyte-derived dendritic cells (mono-DCs). Mycobacterium tuberculosis H37Rv and zymosan preferentially induced IL-23. IL-23 but not IL-12 was efficiently induced by the combination of nucleotide-binding oligodimerization domain and Toll-like receptor (TLR) 2 ligands, which mimics activation by M. tuberculosis, or by the human dectin-1 ligand beta-glucan alone or in combination with TLR2 ligands, mimicking induction by zymosan. TLR2 ligands inhibited IL-12 and increased IL-23 production. DC priming with interferon (IFN) gamma strongly increased IL-12 production, but was not required for IL-23 production and inhibited IL-23 production induced by beta-glucan. The pattern of IL-12 and IL-23 induction was reflected in accumulation of the IL-12p35 and IL-23p19 transcripts, respectively, but not IL-12/23p40. Although IL-23, transforming growth factor beta, and IL-6 contained in the supernatants of activated mono-DCs played a role in the induction of IL-17 by human CD4(+) T cells, IL-1beta, in combination with one or more of those factors, was required for IL-17 production, and its production determined the differential ability of the stimuli used to elicit mono-DCs to produce soluble factors directing IL-17 production. Thus, the differential ability of pathogens to induce antigen-presenting cells to produce cytokines regulates the immune response to infection.
Figures
References
-
- Kobayashi, M., L. Fitz, M. Ryan, R.M. Hewick, S.C. Clark, S. Chan, R. Loudon, F. Sherman, B. Perussia, and G. Trinchieri. 1989. Identification and purification of natural killer cell stimulatory factor (NKSF), a cytokine with multiple biologic effects on human lymphocytes. J. Exp. Med. 170:827–846. - PMC - PubMed
-
- Oppmann, B., R. Lesley, B. Blom, J.C. Timans, Y. Xu, B. Hunte, F. Vega, N. Yu, J. Wang, K. Singh, et al. 2000. Novel p19 protein engages IL-12p40 to form a cytokine, IL-23, with biological activities similar as well as distinct from IL-12. Immunity. 13:715–725. - PubMed
-
- Cooper, A.M., and S.A. Khader. 2007. IL-12p40: an inherently agonistic cytokine. Trends Immunol. 28:33–38. - PubMed
-
- Trinchieri, G. 2003. Interleukin-12 and the regulation of innate resistance and adaptive immunity. Nat. Rev. Immunol. 3:133–146. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
