

An antiinflammatory role for IKKbeta through the inhibition of "classical" macrophage activation
- PMID: 18490491
- PMCID: PMC2413025
- DOI: 10.1084/jem.20080124
An antiinflammatory role for IKKbeta through the inhibition of "classical" macrophage activation
Abstract
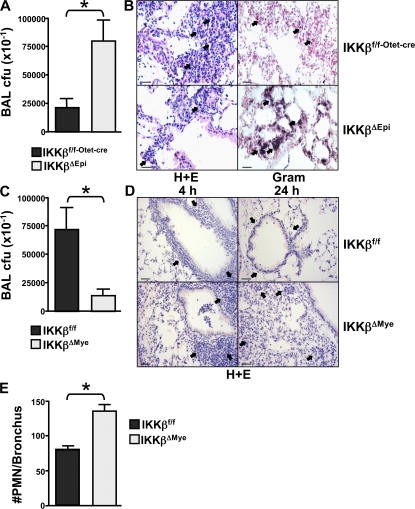
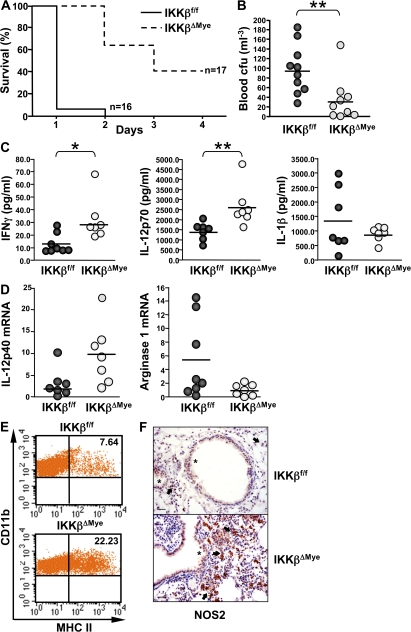
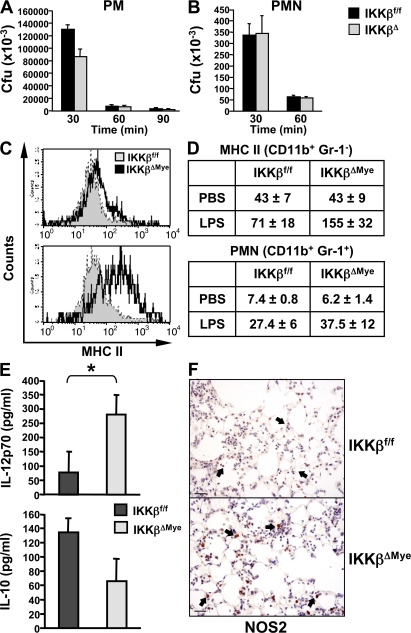
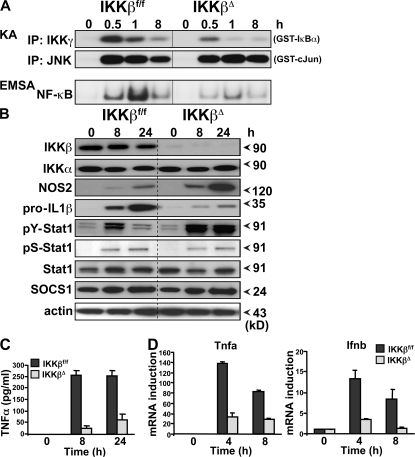
The nuclear factor kappaB (NF-kappaB) pathway plays a central role in inflammation and immunity. In response to proinflammatory cytokines and pathogen-associated molecular patterns, NF-kappaB activation is controlled by IkappaB kinase (IKK)beta. Using Cre/lox-mediated gene targeting of IKKbeta, we have uncovered a tissue-specific role for IKKbeta during infection with group B streptococcus. Although deletion of IKKbeta in airway epithelial cells had the predicted effect of inhibiting inflammation and reducing innate immunity, deletion of IKKbeta in the myeloid lineage unexpectedly conferred resistance to infection that was associated with increased expression of interleukin (IL)-12, inducible nitric oxide synthase (NOS2), and major histocompatibility complex (MHC) class II by macrophages. We also describe a previously unknown role for IKKbeta in the inhibition of signal transducer and activator of transcription (Stat)1 signaling in macrophages, which is critical for IL-12, NOS2, and MHC class II expression. These studies suggest that IKKbeta inhibits the "classically" activated or M1 macrophage phenotype during infection through negative cross talk with the Stat1 pathway. This may represent a mechanism to prevent the over-exuberant activation of macrophages during infection and contribute to the resolution of inflammation. This establishes a new role for IKKbeta in the regulation of macrophage activation with important implications in chronic inflammatory disease, infection, and cancer.
Figures
Comment in
-
IKKbeta/NF-kappaB and the miscreant macrophage.J Exp Med. 2008 Jun 9;205(6):1255-9. doi: 10.1084/jem.20081056. Epub 2008 Jun 2. J Exp Med. 2008. PMID: 18519650 Free PMC article.
References
-
- Karin, M., and Y. Ben-Neriah. 2000. Phosphorylation meets ubiquitination: the control of NF-[kappa]B activity. Annu. Rev. Immunol. 18:621–663. - PubMed
-
- Karin, M., Y. Yamamoto, and Q.M. Wang. 2004. The IKK NF-kappa B system: a treasure trove for drug development. Nat. Rev. Drug Discov. 3:17–26. - PubMed
-
- Lawrence, T., D.W. Gilroy, P.R. Colville-Nash, and D.A. Willoughby. 2001. Possible new role for NF-kappaB in the resolution of inflammation. Nat. Med. 7:1291–1297. - PubMed
-
- Taylor, P.R., and S. Gordon. 2003. Monocyte heterogeneity and innate immunity. Immunity. 19:2–4. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
