

Modulation of TNF-alpha-converting enzyme by the spike protein of SARS-CoV and ACE2 induces TNF-alpha production and facilitates viral entry
- PMID: 18490652
- PMCID: PMC2409424
- DOI: 10.1073/pnas.0711241105
Modulation of TNF-alpha-converting enzyme by the spike protein of SARS-CoV and ACE2 induces TNF-alpha production and facilitates viral entry
Abstract
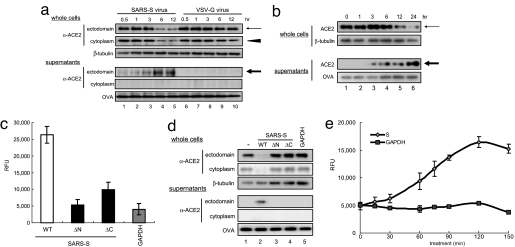
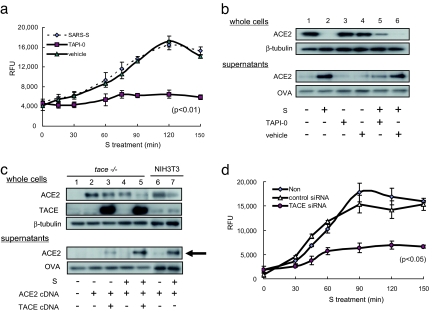
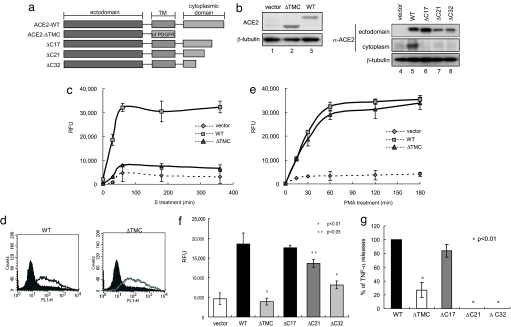
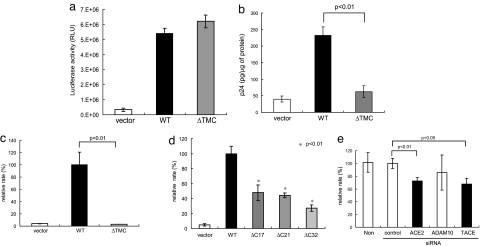
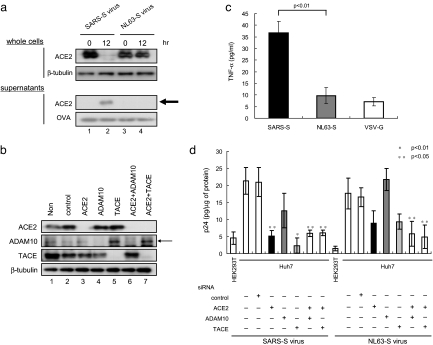
Severe acute respiratory syndrome coronavirus (SARS-CoV) is a high-risk infectious pathogen. In the proposed model of respiratory failure, SARS-CoV down-regulates its receptor, angiotensin-converting enzyme 2 (ACE2), but the mechanism involved is unknown. We found that the spike protein of SARS-CoV (SARS-S) induced TNF-alpha-converting enzyme (TACE)-dependent shedding of the ACE2 ectodomain. The modulation of TACE activity by SARS-S depended on the cytoplasmic domain of ACE2, because deletion mutants of ACE2 lacking the carboxyl-terminal region did not induce ACE2 shedding or TNF-alpha production. In contrast, the spike protein of HNL63-CoV (NL63-S), a CoV that uses ACE2 as a receptor and mainly induces the common cold, caused neither of these cellular responses. Intriguingly, viral infection, judged by real-time RT-PCR analysis of SARS-CoV mRNA expression, was significantly attenuated by deletion of the cytoplasmic tail of ACE2 or knock-down of TACE expression by siRNA. These data suggest that cellular signals triggered by the interaction of SARS-CoV with ACE2 are positively involved in viral entry but lead to tissue damage. These findings may lead to the development of anti-SARS-CoV agents.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Kaiazek TG, et al. A novel coronavirus associated with severe acute respiratory syndrome. N Engl J Med. 2003;15:1953–1966. - PubMed
-
- Tipnis SR, et al. A human homolog of angiotensin-converting enzyme. Cloning and functional expression as a captopril-insensitive carboxypeptidase. J Biol Chem. 2000;275:33238–33243. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
