

Structural basis of actin recognition and arginine ADP-ribosylation by Clostridium perfringens iota-toxin
- PMID: 18490658
- PMCID: PMC2387182
- DOI: 10.1073/pnas.0801215105
Structural basis of actin recognition and arginine ADP-ribosylation by Clostridium perfringens iota-toxin
Abstract
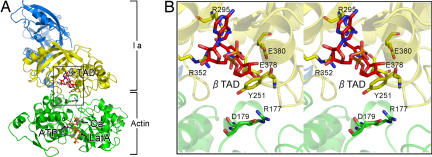
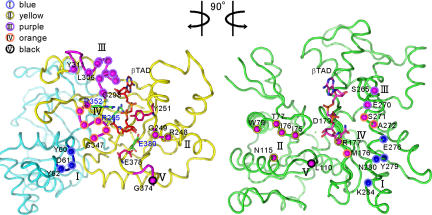
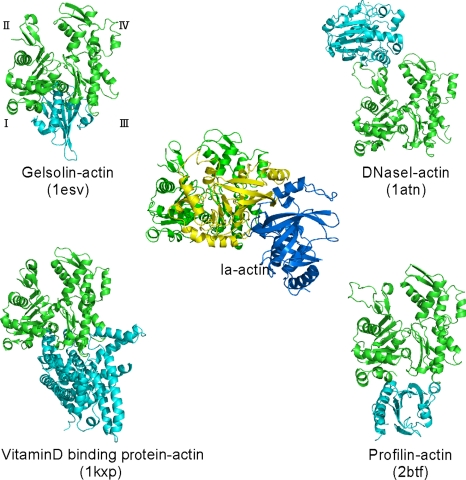
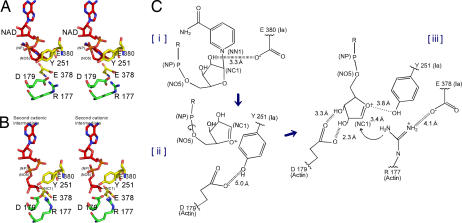
The ADP-ribosylating toxins (ADPRTs) produced by pathogenic bacteria modify intracellular protein and affect eukaryotic cell function. Actin-specific ADPRTs (including Clostridium perfringens iota-toxin and Clostridium botulinum C2 toxin) ADP-ribosylate G-actin at Arg-177, leading to disorganization of the cytoskeleton and cell death. Although the structures of many actin-specific ADPRTs are available, the mechanisms underlying actin recognition and selective ADP-ribosylation of Arg-177 remain unknown. Here we report the crystal structure of actin-Ia in complex with the nonhydrolyzable NAD analog betaTAD at 2.8 A resolution. The structure indicates that Ia recognizes actin via five loops around NAD: loop I (Tyr-60-Tyr-62 in the N domain), loop II (active-site loop), loop III, loop IV (PN loop), and loop V (ADP-ribosylating turn-turn loop). We used site-directed mutagenesis to confirm that loop I on the N domain and loop II are essential for the ADP-ribosyltransferase activity. Furthermore, we revealed that Glu-378 on the EXE loop is in close proximity to Arg-177 in actin, and we proposed that the ADP-ribosylation of Arg-177 proceeds by an SN1 reaction via first an oxocarbenium ion intermediate and second a cationic intermediate by alleviating the strained conformation of the first oxocarbenium ion. Our results suggest a common reaction mechanism for ADPRTs. Moreover, the structure might be of use in rational drug design to block toxin-substrate recognition.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
Similar articles
-
Arginine ADP-ribosylation mechanism based on structural snapshots of iota-toxin and actin complex.Proc Natl Acad Sci U S A. 2013 Mar 12;110(11):4267-72. doi: 10.1073/pnas.1217227110. Epub 2013 Feb 4. Proc Natl Acad Sci U S A. 2013. PMID: 23382240 Free PMC article.
-
Crystal structure and site-directed mutagenesis of enzymatic components from Clostridium perfringens iota-toxin.J Mol Biol. 2003 Jan 17;325(3):471-83. doi: 10.1016/s0022-2836(02)01247-0. J Mol Biol. 2003. PMID: 12498797
-
Crystal structure and novel recognition motif of rho ADP-ribosylating C3 exoenzyme from Clostridium botulinum: structural insights for recognition specificity and catalysis.J Mol Biol. 2001 Jan 5;305(1):95-107. doi: 10.1006/jmbi.2000.4292. J Mol Biol. 2001. PMID: 11114250
-
Clostridial ADP-ribosylating toxins: effects on ATP and GTP-binding proteins.Mol Cell Biochem. 1994 Sep;138(1-2):167-76. doi: 10.1007/BF00928459. Mol Cell Biochem. 1994. PMID: 7898461 Review.
-
Bidirectional attack on the actin cytoskeleton. Bacterial protein toxins causing polymerization or depolymerization of actin.Toxicon. 2012 Sep 15;60(4):572-81. doi: 10.1016/j.toxicon.2012.04.338. Epub 2012 Apr 19. Toxicon. 2012. PMID: 22543189 Review.
Cited by
-
NMR solution structures of Runella slithyformis RNA 2'-phosphotransferase Tpt1 provide insights into NAD+ binding and specificity.Nucleic Acids Res. 2021 Sep 27;49(17):9607-9624. doi: 10.1093/nar/gkab241. Nucleic Acids Res. 2021. PMID: 33880546 Free PMC article.
-
Cryo-EM structures of the translocational binary toxin complex CDTa-bound CDTb-pore from Clostridioides difficile.Nat Commun. 2022 Oct 17;13(1):6119. doi: 10.1038/s41467-022-33888-4. Nat Commun. 2022. PMID: 36253419 Free PMC article.
-
A type VI secretion system effector protein, VgrG1, from Aeromonas hydrophila that induces host cell toxicity by ADP ribosylation of actin.J Bacteriol. 2010 Jan;192(1):155-68. doi: 10.1128/JB.01260-09. J Bacteriol. 2010. PMID: 19880608 Free PMC article.
-
Characterization of an actin-targeting ADP-ribosyltransferase from Aeromonas hydrophila.J Biol Chem. 2012 Oct 26;287(44):37030-41. doi: 10.1074/jbc.M112.397612. Epub 2012 Sep 11. J Biol Chem. 2012. PMID: 22969084 Free PMC article.
-
Arginine ADP-ribosylation mechanism based on structural snapshots of iota-toxin and actin complex.Proc Natl Acad Sci U S A. 2013 Mar 12;110(11):4267-72. doi: 10.1073/pnas.1217227110. Epub 2013 Feb 4. Proc Natl Acad Sci U S A. 2013. PMID: 23382240 Free PMC article.
References
-
- Holbourn KP, Shone CC, Acharya KR. A family of killer toxins. Exploring the mechanism of ADP-ribosylating toxins. FEBS J. 2006;273:4579–4593. - PubMed
-
- Moss J, Garrison S, Oppenheimer NJ, Richardson SH. NAD-dependent ADP-ribosylation of arginine and proteins by Escherichia coli heat-labile enterotoxin. J Biol Chem. 1979;254:6270–6272. - PubMed
-
- Van Ness BG, Howard JB, Bodley JW. ADP-ribosylation of elongation factor 2 by diphtheria toxin. Isolation and properties of the novel ribosyl-amino acid and its hydrolysis products. J Biol Chem. 1980;255:10717–10720. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
