

CD28/CD154 blockade prevents autoimmune diabetes by inducing nondeletional tolerance after effector t-cell inhibition and regulatory T-cell expansion
- PMID: 18492787
- PMCID: PMC2551677
- DOI: 10.2337/db07-1712
CD28/CD154 blockade prevents autoimmune diabetes by inducing nondeletional tolerance after effector t-cell inhibition and regulatory T-cell expansion
Abstract
Objective: Blocking T-cell signaling is an effective means to prevent autoimmunity and allograft rejection in many animal models, yet the clinical translation of many of these approaches has not resulted in the success witnessed in experimental systems. Improved understanding of these approaches may assist in developing safe and effective means to treat disorders such as autoimmune diabetes.
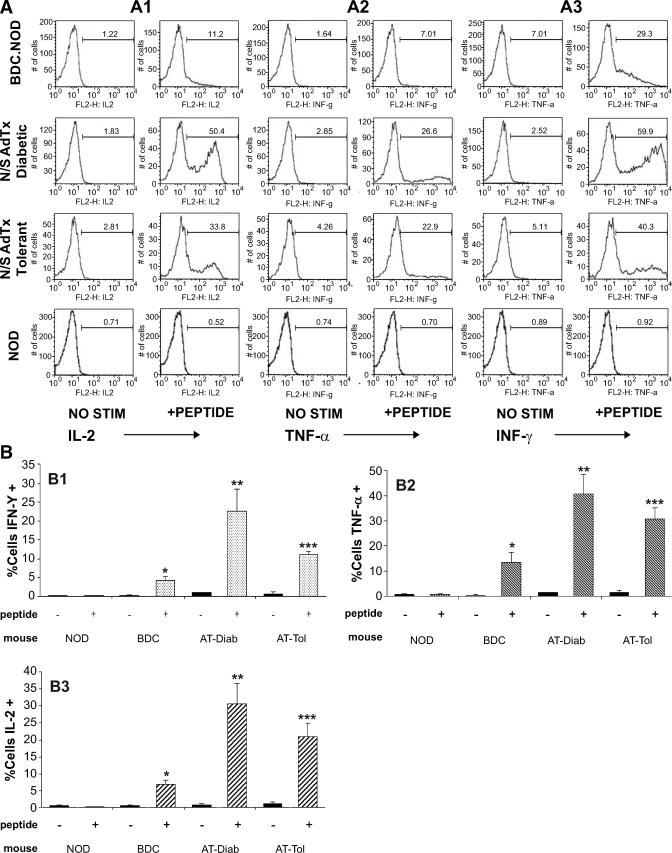
Research design and methods: We studied the effect of anti-CD154 and CTLA4-Ig on diabetes development, and the requirements to induce tolerance in nod.scid mice after transfer of transgenic beta-cell reactive BDC2.5.NOD T-cells.
Results: Nod.scid recipients of diabetogenic BDC2.5.NOD cells were protected indefinitely from diabetes by a short course of combined costimulation blockade, despite the continued diabetogenic potential of their T-cells. The presence of pathogenic T-cells in the absence of disease indicates peripheral immune tolerance. T-cell maturation occurred in protected recipients, yet costimulation blockade temporarily blunted early T-cell proliferation in draining pancreatic nodes. Tolerance required preexisting regulatory T-cells (Tregs), and protected recipients had greater numbers of Tregs than diabetic recipients. Diabetes protection was successful in the presence of homeostatic expansion and high T-cell precursor frequency, both obstacles to tolerance induction in other models of antigen-specific immunity.
Conclusions: Immunotherapies that selectively suppress effector T-cells while permitting the development of natural regulatory mechanisms may have a unique role in establishing targeted long-standing immune protection and peripheral tolerance. Understanding the mechanism of these approaches may assist in the design and use of therapies for human conditions, such as type 1 diabetes.
Figures









Similar articles
-
Islet allograft rejection in nonobese diabetic mice involves the common gamma-chain and CD28/CD154-dependent and -independent mechanisms.J Immunol. 2003 Oct 1;171(7):3878-85. doi: 10.4049/jimmunol.171.7.3878. J Immunol. 2003. PMID: 14500690
-
CD8+CD122+PD-1+ Tregs Synergize With Costimulatory Blockade of CD40/CD154, but Not B7/CD28, to Prolong Murine Allograft Survival.Front Immunol. 2019 Feb 26;10:306. doi: 10.3389/fimmu.2019.00306. eCollection 2019. Front Immunol. 2019. PMID: 30863408 Free PMC article.
-
Costimulation blockade of both inducible costimulator and CD40 ligand induces dominant tolerance to islet allografts and prevents spontaneous autoimmune diabetes in the NOD mouse.Diabetes. 2006 Jan;55(1):27-33. Diabetes. 2006. PMID: 16380473
-
Control of type 1 autoimmune diabetes by naturally occurring CD4+CD25+ regulatory T lymphocytes in neonatal NOD mice.Ann N Y Acad Sci. 2005 Jun;1051:72-87. doi: 10.1196/annals.1361.048. Ann N Y Acad Sci. 2005. PMID: 16126946 Review.
-
Therapeutic vaccination using CD4+CD25+ antigen-specific regulatory T cells.Proc Natl Acad Sci U S A. 2004 Oct 5;101 Suppl 2(Suppl 2):14622-6. doi: 10.1073/pnas.0405234101. Epub 2004 Aug 20. Proc Natl Acad Sci U S A. 2004. PMID: 15322272 Free PMC article. Review.
Cited by
-
Targeting co-stimulatory pathways: transplantation and autoimmunity.Nat Rev Nephrol. 2014 Jan;10(1):14-24. doi: 10.1038/nrneph.2013.183. Epub 2013 Oct 8. Nat Rev Nephrol. 2014. PMID: 24100403 Free PMC article. Review.
-
Endogenous memory CD8 T cells are activated within cardiac allografts without mediating rejection.Am J Transplant. 2013 Sep;13(9):2293-307. doi: 10.1111/ajt.12372. Epub 2013 Aug 5. Am J Transplant. 2013. PMID: 23914930 Free PMC article.
-
Central memory CD8+ T lymphocytes mediate lung allograft acceptance.J Clin Invest. 2014 Mar;124(3):1130-43. doi: 10.1172/JCI71359. Epub 2014 Feb 24. J Clin Invest. 2014. PMID: 24569377 Free PMC article.
-
Of the multiple mechanisms leading to type 1 diabetes, T cell receptor revision may play a prominent role (is type 1 diabetes more than a single disease?).Clin Exp Immunol. 2016 Sep;185(3):271-80. doi: 10.1111/cei.12819. Epub 2016 Jul 25. Clin Exp Immunol. 2016. PMID: 27271348 Free PMC article. Review.
-
Distinct peripheral blood molecular signature emerges with successful tacrolimus withdrawal in kidney transplant recipients.Am J Transplant. 2020 Dec;20(12):3477-3485. doi: 10.1111/ajt.15979. Epub 2020 May 27. Am J Transplant. 2020. PMID: 32459070 Free PMC article. Clinical Trial.
References
-
- Larsen CP, Elwood ET, Alexander DZ, Ritchie SC, Hendrix R, Tucker-Burden C, Cho HR, Aruffo A, Hollenbaugh D, Linsley PS, Winn KJ, Pearson TC: Long-term acceptance of skin and cardiac allografts after blocking CD40 and CD28 pathways. Nature 381:434–438, 1996 - PubMed
-
- Akalin E, Chandraker A, Russell ME, Turka LA, Hancock WW, Sayegh MH: CD28–B7 T cell costimulatory blockade by CTLA4Ig in the rat renal allograft model: inhibition of cell-mediated and humoral immune responses in vivo. Transplantation 62:1942–1945, 1996 - PubMed
-
- Adams AB, Shirasugi N, Durham MM, Strobert E, Anderson D, Rees P, Cowan S, Xu H, Blinder Y, Cheung M, Hollenbaugh D, Kenyon NS, Pearson TC, Larsen CP: Calcineurin inhibitor-free CD28 blockade-based protocol protects allogeneic islets in nonhuman primates. Diabetes 51:265–270, 2002 - PubMed
-
- Li Y, Li XC, Zheng XX, Wells AD, Turka LA, Strom TB: Blocking both signal 1 and signal 2 of T-cell activation prevents apoptosis of alloreactive T cells and induction of peripheral allograft tolerance. Nat Med 5:1298–1302, 1999 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
