

A gene duplication affecting expression of the ovine ASIP gene is responsible for white and black sheep
- PMID: 18493018
- PMCID: PMC2493430
- DOI: 10.1101/gr.072090.107
A gene duplication affecting expression of the ovine ASIP gene is responsible for white and black sheep
Abstract
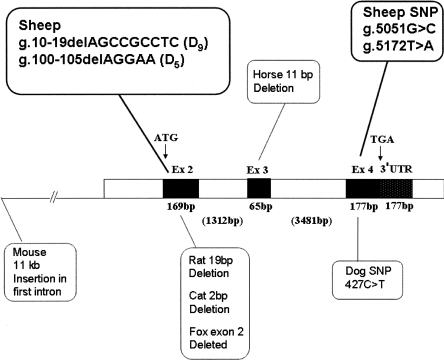
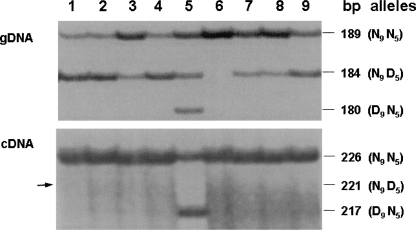
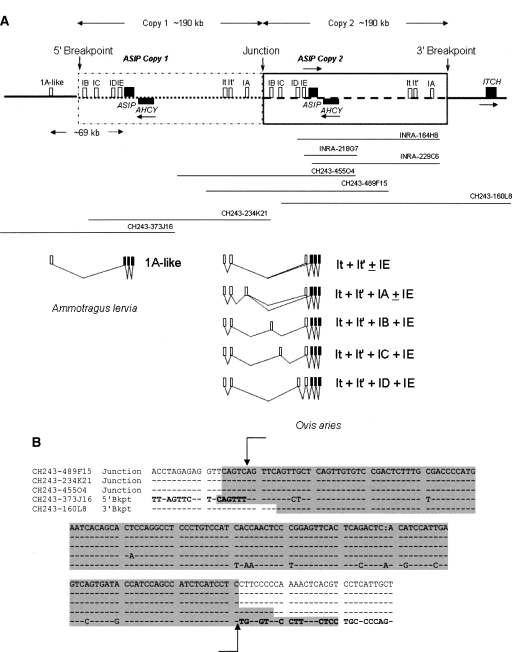
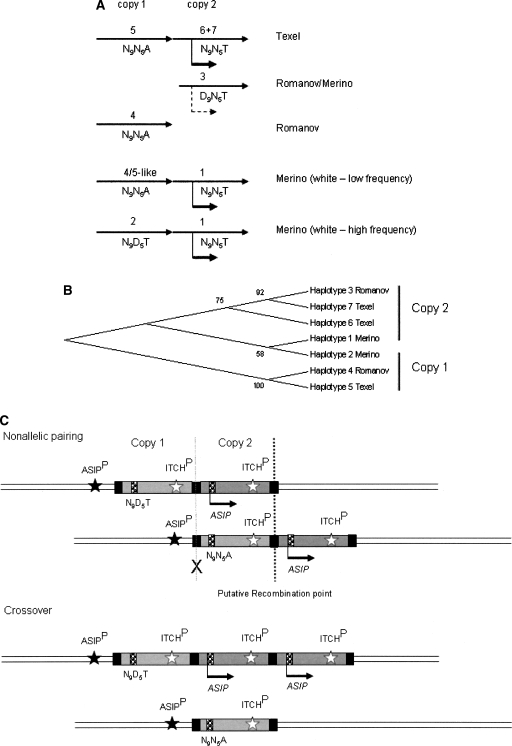
Agouti signaling protein (ASIP) functions to regulate pigmentation in mice, while its role in many other animals and in humans has not been fully determined. In this study, we identify a 190-kb tandem duplication encompassing the ovine ASIP and AHCY coding regions and the ITCH promoter region as the genetic cause of white coat color of dominant white/tan (A(Wt)) agouti sheep. The duplication 5' breakpoint is located upstream of the ASIP coding sequence. Ubiquitous expression of a second copy of the ASIP coding sequence regulated by a duplicated copy of the nearby ITCH promoter causes the white sheep phenotype. A single copy ASIP gene with a silenced ASIP promoter occurs in recessive black sheep. In contrast, a single copy functional wild-type (A(+)) ASIP is responsible for the ancient Barbary sheep coat color phenotype. The gene duplication was facilitated by homologous recombination between two non-LTR SINE sequences flanking the duplicated segment. This is the first sheep trait attributable to gene duplication and shows nonallelic homologous recombination and gene conversion events at the ovine ASIP locus could have an important role in the evolution of sheep pigmentation.
Figures


Similar articles
-
Analysis of agouti signaling protein (ASIP) gene polymorphisms and association with coat color in Tibetan sheep (Ovis aries).Genet Mol Res. 2015 Feb 6;14(1):1200-9. doi: 10.4238/2015.February.6.22. Genet Mol Res. 2015. PMID: 25730058
-
Epistatic Interaction of the Melanocortin 1 Receptor and Agouti Signaling Protein Genes Modulates Wool Color in the Brazilian Creole Sheep.J Hered. 2016 Nov;107(6):544-52. doi: 10.1093/jhered/esw037. Epub 2016 Jun 9. J Hered. 2016. PMID: 27288530
-
Differences in the expression of the ASIP gene are involved in the recessive black coat colour pattern in sheep: evidence from the rare Xalda sheep breed.Anim Genet. 2008 Jun;39(3):290-3. doi: 10.1111/j.1365-2052.2008.01712.x. Epub 2008 Mar 31. Anim Genet. 2008. PMID: 18384465
-
Agouti: from mouse to man, from skin to fat.Pigment Cell Res. 2002 Feb;15(1):10-8. doi: 10.1034/j.1600-0749.2002.00039.x. Pigment Cell Res. 2002. PMID: 11837451 Review.
-
The genetics of colour in fat-tailed sheep: a review.Trop Anim Health Prod. 2011 Oct;43(7):1245-65. doi: 10.1007/s11250-011-9850-0. Epub 2011 May 5. Trop Anim Health Prod. 2011. PMID: 21544704 Review.
Cited by
-
Variants at the ASIP locus contribute to coat color darkening in Nellore cattle.Genet Sel Evol. 2021 Apr 28;53(1):40. doi: 10.1186/s12711-021-00633-2. Genet Sel Evol. 2021. PMID: 33910501 Free PMC article.
-
Genome-Wide Detection of CNVs and Their Association with Meat Tenderness in Nelore Cattle.PLoS One. 2016 Jun 27;11(6):e0157711. doi: 10.1371/journal.pone.0157711. eCollection 2016. PLoS One. 2016. PMID: 27348523 Free PMC article.
-
Genomic Structural Diversity in Local Goats: Analysis of Copy-Number Variations.Animals (Basel). 2020 Jun 16;10(6):1040. doi: 10.3390/ani10061040. Animals (Basel). 2020. PMID: 32560248 Free PMC article.
-
Genetic Basis and Evolution of Structural Color Polymorphism in an Australian Songbird.Mol Biol Evol. 2024 Mar 1;41(3):msae046. doi: 10.1093/molbev/msae046. Mol Biol Evol. 2024. PMID: 38415852 Free PMC article.
-
Genome-Wide Specific Selection in Three Domestic Sheep Breeds.PLoS One. 2015 Jun 17;10(6):e0128688. doi: 10.1371/journal.pone.0128688. eCollection 2015. PLoS One. 2015. PMID: 26083354 Free PMC article.
References
-
- Adalsteinsson S. Colour inheritance in Icelandic sheep and relation between colour, fertility and fertilisation. J. Agric. Res. Iceland. 1970;2:3–135.
-
- Bennett D.C., Lamoreux M.L., Lamoreux M.L. The colour loci of mice—A genetic century. Pigment Cell Res. 2003;16:333–344. - PubMed
-
- Bischof J.M., Chiang A.R., Scheetz T.E., Stone E.M., Casavant T.L., Sheffield V.C., Braun T.A., Chiang A.R., Scheetz T.E., Stone E.M., Casavant T.L., Sheffield V.C., Braun T.A., Scheetz T.E., Stone E.M., Casavant T.L., Sheffield V.C., Braun T.A., Stone E.M., Casavant T.L., Sheffield V.C., Braun T.A., Casavant T.L., Sheffield V.C., Braun T.A., Sheffield V.C., Braun T.A., Braun T.A. Genome-wide identification of pseudogenes capable of disease-causing gene conversion. Hum. Mutat. 2006;27:545–552. - PubMed
-
- Bonilla C., Boxill L.A., Donald S.A., Williams T., Sylvester N., Parra E.J., Dios S., Norton H.L., Shriver M.D., Kittles R.A., Boxill L.A., Donald S.A., Williams T., Sylvester N., Parra E.J., Dios S., Norton H.L., Shriver M.D., Kittles R.A., Donald S.A., Williams T., Sylvester N., Parra E.J., Dios S., Norton H.L., Shriver M.D., Kittles R.A., Williams T., Sylvester N., Parra E.J., Dios S., Norton H.L., Shriver M.D., Kittles R.A., Sylvester N., Parra E.J., Dios S., Norton H.L., Shriver M.D., Kittles R.A., Parra E.J., Dios S., Norton H.L., Shriver M.D., Kittles R.A., Dios S., Norton H.L., Shriver M.D., Kittles R.A., Norton H.L., Shriver M.D., Kittles R.A., Shriver M.D., Kittles R.A., Kittles R.A. The 8818G allele of the agouti signaling protein (ASIP) gene is ancestral and is associated with darker skin colour in African Americans. Hum. Genet. 2005;116:402–406. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials