

Nucleotide diversity in the mitochondrial and nuclear compartments of Chlamydomonas reinhardtii: investigating the origins of genome architecture
- PMID: 18495022
- PMCID: PMC2412866
- DOI: 10.1186/1471-2148-8-156
Nucleotide diversity in the mitochondrial and nuclear compartments of Chlamydomonas reinhardtii: investigating the origins of genome architecture
Abstract
Background: The magnitude of intronic and intergenic DNA can vary substantially both within and among evolutionary lineages; however, the forces responsible for this disparity in genome compactness are conjectural. One explanation, termed the mutational-burden hypothesis, posits that genome compactness is primarily driven by two nonadaptive processes: mutation and random genetic drift - the effects of which can be discerned by measuring the nucleotide diversity at silent sites (pisilent), defined as noncoding sites and the synonymous sites of protein-coding regions. The mutational-burden hypothesis holds that pisilent is negatively correlated to genome compactness. We used the model organism Chlamydomonas reinhardtii, which has a streamlined, coding-dense mitochondrial genome and an noncompact, intron-rich nuclear genome, to investigate the mutational-burden hypothesis. For measuring pisilent we sequenced the complete mitochondrial genome and portions of 7 nuclear genes from 7 geographical isolates of C. reinhardtii.
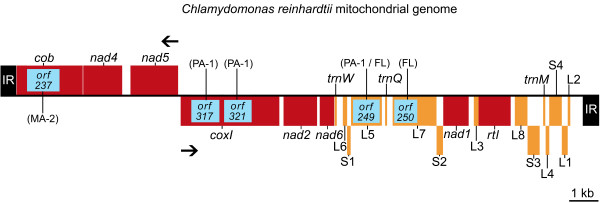
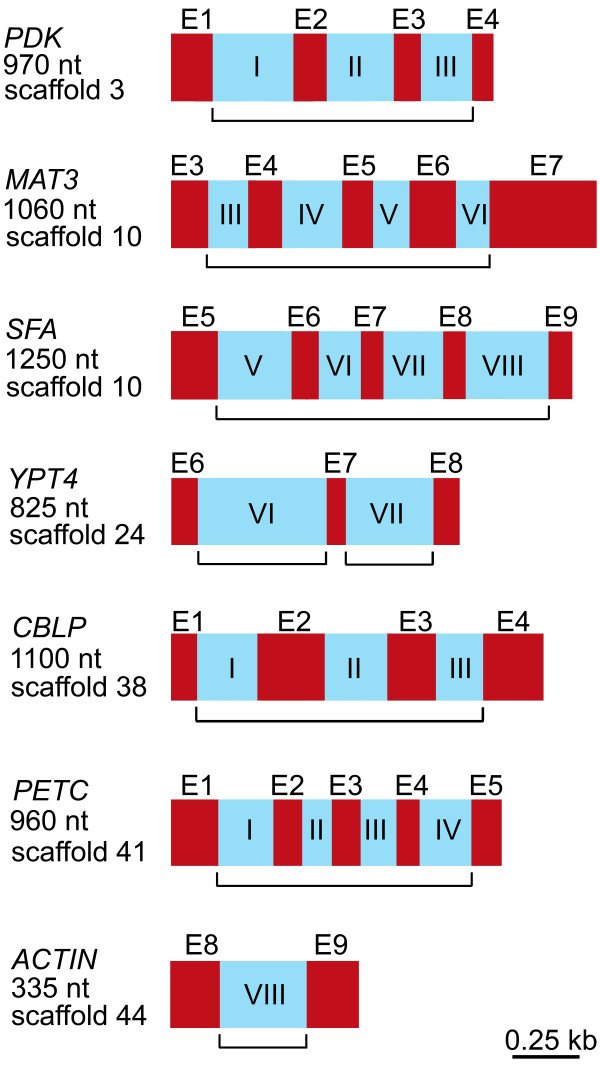
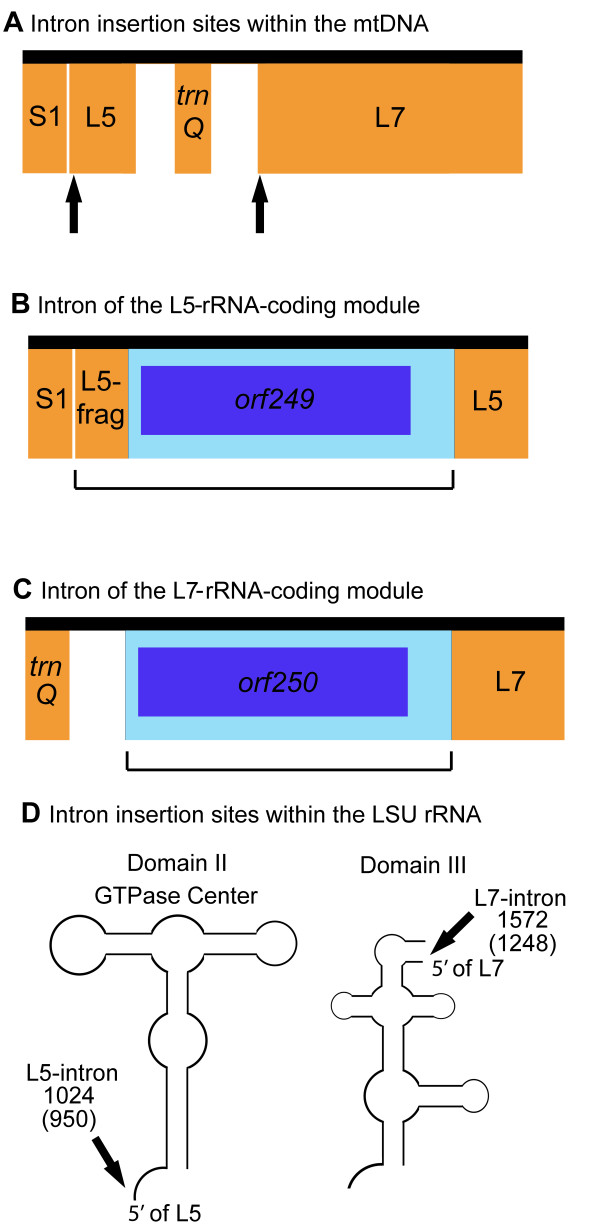
Results: We found significantly more nucleotide diversity in the nuclear compartment of C. reinhardtii than in the mitochondrial compartment: net values of pisilent for the nuclear and mitochondrial genomes were 32 x 10-3 and 8.5 x 10-3, respectively; and when insertions and deletions (indels) are factored in, these values become 49 x 10-3 for the nuclear DNA and 11 x 10-3 for the mitochondrial DNA (mtDNA). Furthermore, our investigations of C. reinhardtii revealed 4 previously undiscovered mitochondrial introns, one of which contains a fragment of the large-subunit (LSU) rRNA gene and another of which is found in a region of the LSU-rRNA gene not previously reported (for any taxon) to contain introns.
Conclusion: At first glance our results are in opposition to the mutational-burden hypothesis: pisilent was approximately 4 times greater in the nuclear compartment of C. reinhardtii relative to the mitochondrial compartment. However, when we consider the encumbrance of noncoding DNA in each of these C. reinhardtii compartments, we conclude that introns in the mtDNA impose a greater burden than those in the nuclear DNA and suggest that the same may be true for the intergenic regions. Overall, we cannot reject the mutational-burden hypothesis and feel that more data on nucleotide diversity from green algae and other protists are needed.
Figures
References
-
- Lynch M. The Origins of Genome Architecture. Sunderland: Sinauer Associates, Inc; 2007.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
