

The cAMP-activated GTP exchange factor, Epac1 upregulates plasma membrane and nuclear Akt kinase activities in 8-CPT-2-O-Me-cAMP-stimulated macrophages: Gene silencing of the cAMP-activated GTP exchange Epac1 prevents 8-CPT-2-O-Me-cAMP activation of Akt activity in macrophages
- PMID: 18495429
- PMCID: PMC2519236
- DOI: 10.1016/j.cellsig.2008.04.002
The cAMP-activated GTP exchange factor, Epac1 upregulates plasma membrane and nuclear Akt kinase activities in 8-CPT-2-O-Me-cAMP-stimulated macrophages: Gene silencing of the cAMP-activated GTP exchange Epac1 prevents 8-CPT-2-O-Me-cAMP activation of Akt activity in macrophages
Abstract
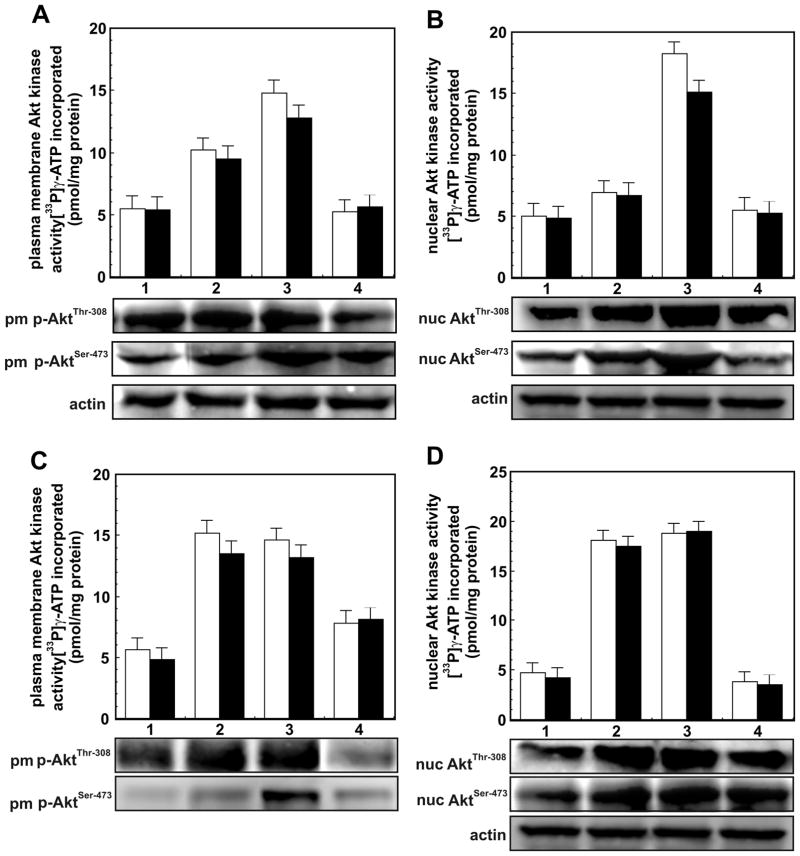
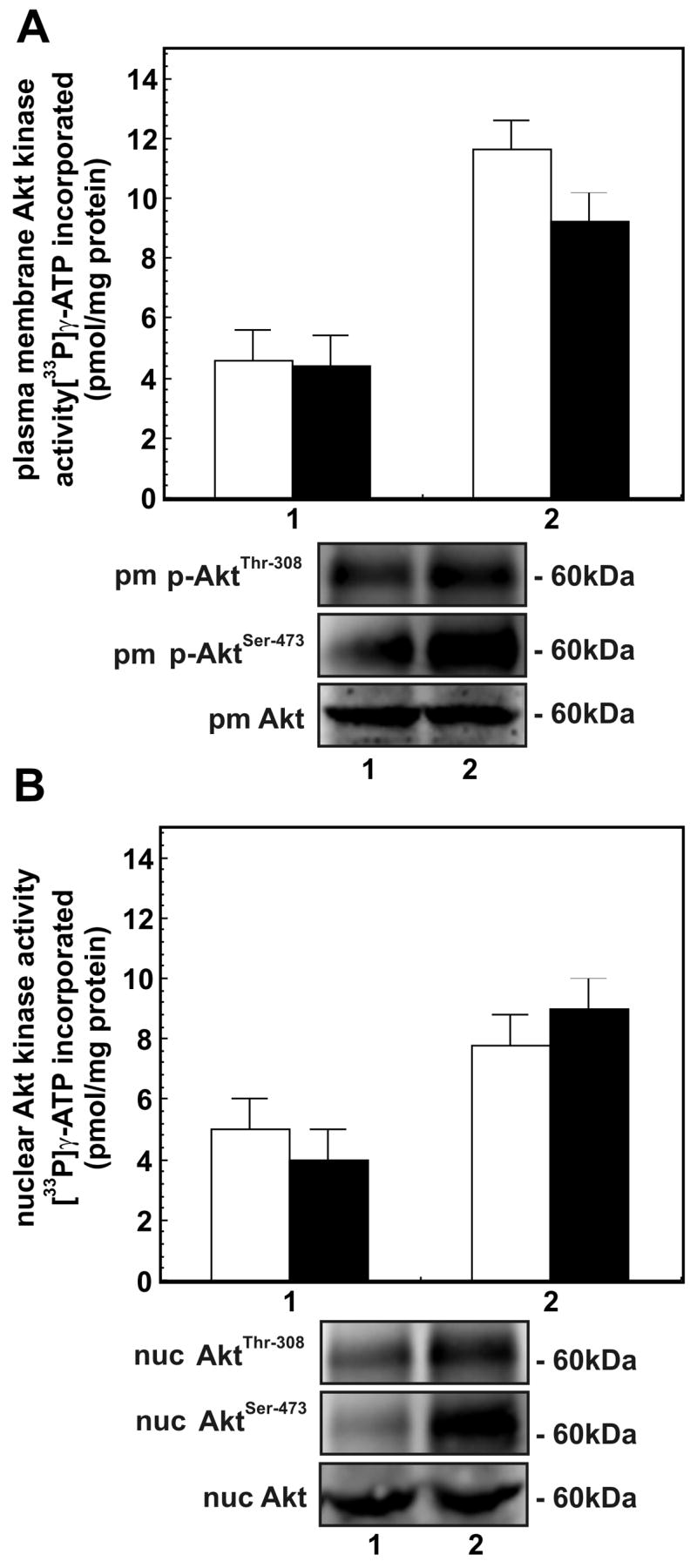
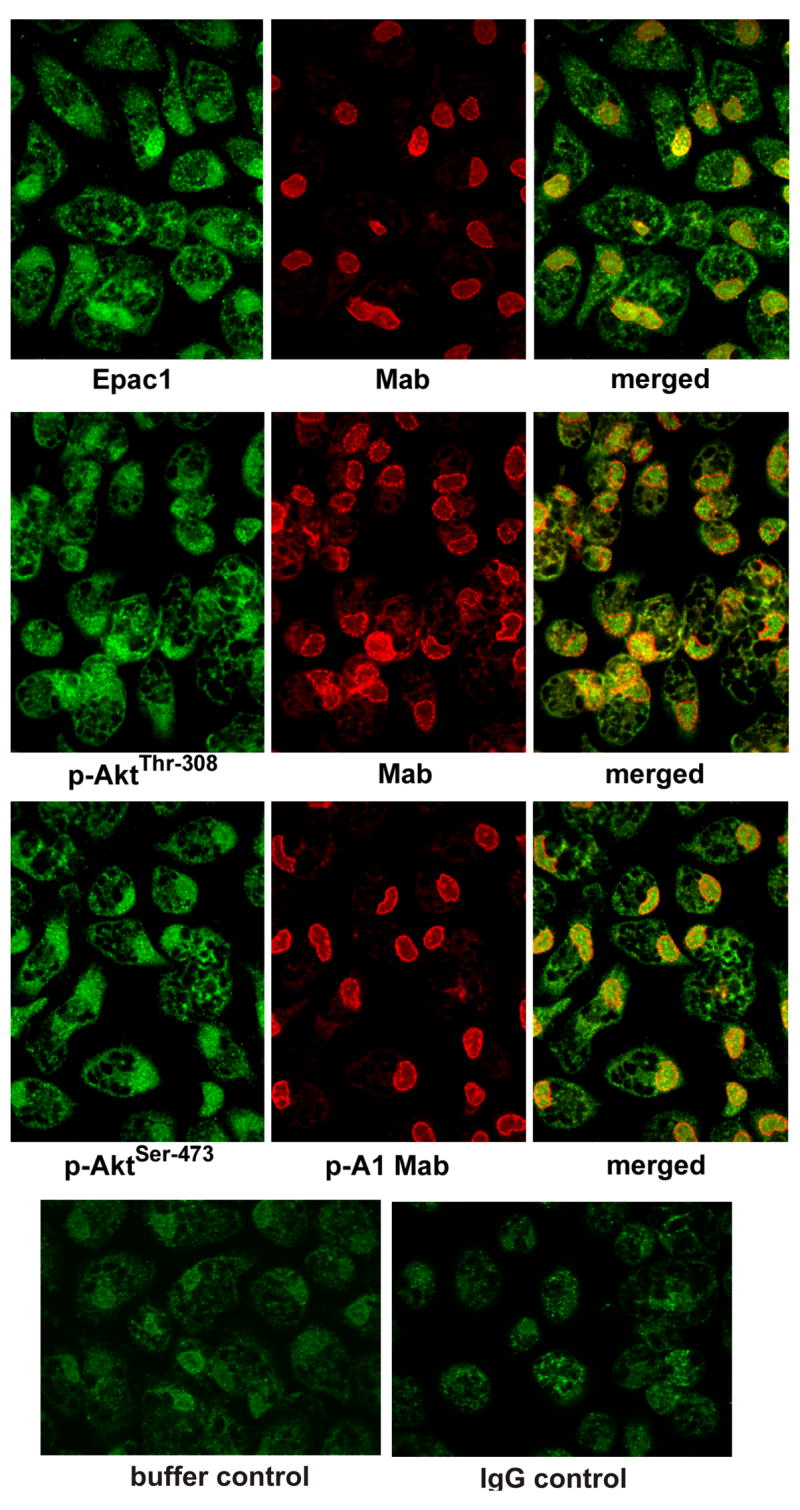
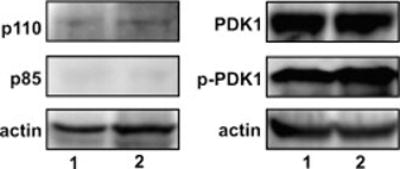
cAMP regulates a wide range of processes through its downstream effectors including PKA, and the family of guanine nucleotide exchange factors. Depending on the cell type, cAMP inhibits or stimulates growth and proliferation in a PKA-dependent or independent manner. PKA-independent effects are mediated by PI 3-kinases-Akt signaling and EPAC1 (exchange protein directly activated by cAMP) activation. Recently, we reported PKA-independent activation of the protein kinase Akt as well co-immunoprecipitation of Epac1 with Rap1, p-Akt(Thr-308), and p-Akt(Ser-473) in forskolin-stimulated macrophages. To further probe the role of Epac1 in Akt protein kinase activation and cellular proliferation, we employed the cAMP analog 8-CPT-2-O-Me-cAMP, which selectively binds to Epac1 and triggers Epac1 signaling. We show the association of Epac1 with activated Akt kinases by co-immunoprecipitation and GST-pulldown assays. Silencing Epac1 gene expression by RNA interference significantly reduced levels of Epac1 mRNA, Epac protein, Rap1 GTP, p-ERK1/2, p-B-Raf, p110alpha catalytic subunit of PI 3-kinase, p-PDK, and p-p(70s6k). Silencing Epac1 gene expression by RNA interference also suppressed 8-CPT-2-O-Me-cAMP-upregulated protein and DNA synthesis. Concomitantly, 8-CPT-2-O-Me-cAMP-mediated upregulation of Akt(Thr-308) protein kinase activity and p-Akt(Thr-308) levels was prevented in plasma membranes and nuclei of the cells. In contrast, silencing Epac1 gene expression reduced Akt(Ser-473) kinase activity and p-Akt(Ser-473) levels in plasma membranes, but showed negligible effects on nuclear activity. In conclusion, we show that cAMP-induced Akt kinase activation and cellular proliferation is mediated by Epac1 which appears to function as an accessory protein for Akt activation.
Figures




Similar articles
-
Interaction between TCL1 and Epac1 in the activation of Akt kinases in plasma membranes and nuclei of 8-CPT-2-O-Me-cAMP-stimulated macrophages.Cell Signal. 2008 Jan;20(1):130-8. doi: 10.1016/j.cellsig.2007.10.008. Epub 2007 Oct 12. Cell Signal. 2008. PMID: 17993260 Free PMC article.
-
Coordinate regulation of forskolin-induced cellular proliferation in macrophages by protein kinase A/cAMP-response element-binding protein (CREB) and Epac1-Rap1 signaling: effects of silencing CREB gene expression on Akt activation.J Biol Chem. 2005 Nov 18;280(46):38276-89. doi: 10.1074/jbc.M507332200. Epub 2005 Sep 19. J Biol Chem. 2005. PMID: 16172130
-
Upregulation of mTORC2 activation by the selective agonist of EPAC, 8-CPT-2Me-cAMP, in prostate cancer cells: assembly of a multiprotein signaling complex.J Cell Biochem. 2012 May;113(5):1488-500. doi: 10.1002/jcb.24018. J Cell Biochem. 2012. Retraction in: J Cell Biochem. 2025 Jul;126(7):e70048. doi: 10.1002/jcb.70048. PMID: 22173835 Retracted.
-
Epac-selective cAMP analogs: new tools with which to evaluate the signal transduction properties of cAMP-regulated guanine nucleotide exchange factors.Cell Signal. 2008 Jan;20(1):10-20. doi: 10.1016/j.cellsig.2007.07.009. Epub 2007 Jul 25. Cell Signal. 2008. PMID: 17716863 Free PMC article. Review.
-
The future of EPAC-targeted therapies: agonism versus antagonism.Trends Pharmacol Sci. 2015 Apr;36(4):203-14. doi: 10.1016/j.tips.2015.02.003. Epub 2015 Mar 3. Trends Pharmacol Sci. 2015. PMID: 25744542 Free PMC article. Review.
Cited by
-
cAMP-GEF cytoprotection by Src tyrosine kinase activation of phosphoinositide-3-kinase p110 beta/alpha in rat hepatocytes.Am J Physiol Gastrointest Liver Physiol. 2009 Apr;296(4):G764-74. doi: 10.1152/ajpgi.90622.2008. Epub 2009 Feb 5. Am J Physiol Gastrointest Liver Physiol. 2009. PMID: 19196950 Free PMC article.
-
RIAM (Rap1-interacting adaptor molecule) regulates complement-dependent phagocytosis.Cell Mol Life Sci. 2013 Jul;70(13):2395-410. doi: 10.1007/s00018-013-1268-6. Epub 2013 Feb 19. Cell Mol Life Sci. 2013. PMID: 23420480 Free PMC article.
-
Transcriptional modulation by VIP: a rational target against inflammatory disease.Clin Epigenetics. 2011 Aug;2(2):213-22. doi: 10.1007/s13148-011-0036-4. Epub 2011 May 18. Clin Epigenetics. 2011. PMID: 22704338 Free PMC article.
-
Multiple facets of cAMP signalling and physiological impact: cAMP compartmentalization in the lung.Pharmaceuticals (Basel). 2012 Nov 30;5(12):1291-331. doi: 10.3390/ph5121291. Pharmaceuticals (Basel). 2012. PMID: 24281338 Free PMC article.
-
cAMP-guanine exchange factor protection from bile acid-induced hepatocyte apoptosis involves glycogen synthase kinase regulation of c-Jun NH2-terminal kinase.Am J Physiol Gastrointest Liver Physiol. 2011 Aug;301(2):G385-400. doi: 10.1152/ajpgi.00430.2010. Epub 2011 May 5. Am J Physiol Gastrointest Liver Physiol. 2011. PMID: 21546580 Free PMC article.
References
-
- Datta SR, Brunet A, Greenberg MA. Genes Dev. 1999;13:2905–2927. - PubMed
-
- Brazil DP, Yang ZZ, Hemmings BA. Trends Biochem Sci. 2004;29:233–242. - PubMed
-
- Bellacosa A, Kumar cC, DiCristofano AD, Testa JR. Adv Cancer Res. 2005;94:29–86. - PubMed
-
- Woodgett JR. Curr Opin Cell Biol. 2005;17:150–157. - PubMed
-
- Du K, Tsichlis PN. Oncogene. 2005;24:7401–7409. - PubMed
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous
