

Agouti-related peptide and MC3/4 receptor agonists both inhibit excitatory hypothalamic ventromedial nucleus neurons
- PMID: 18495877
- PMCID: PMC2634774
- DOI: 10.1523/JNEUROSCI.0749-08.2008
Agouti-related peptide and MC3/4 receptor agonists both inhibit excitatory hypothalamic ventromedial nucleus neurons
Abstract
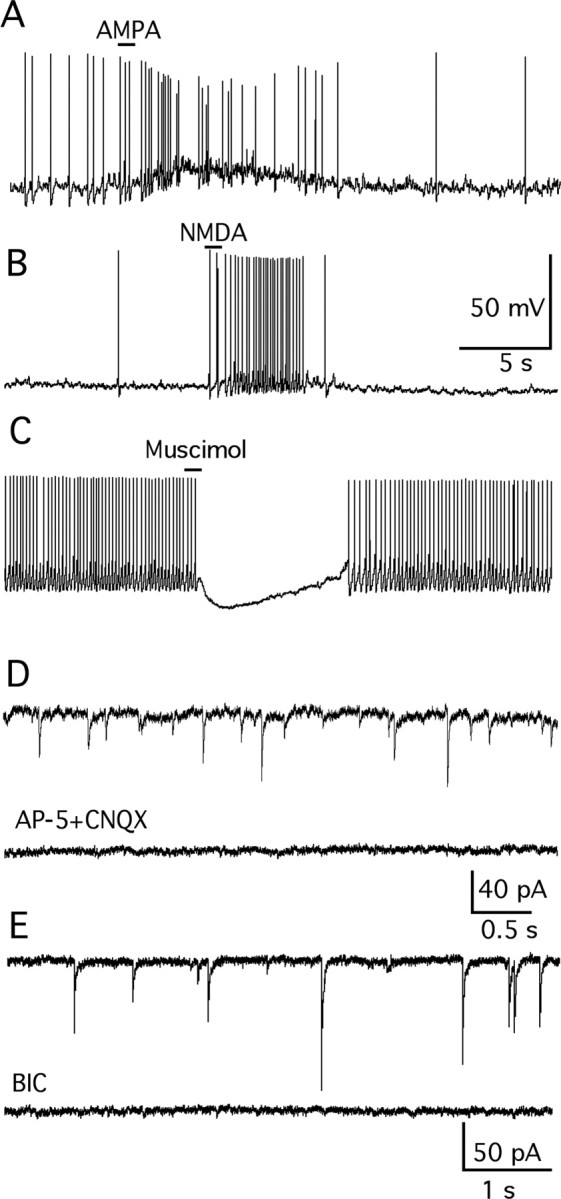
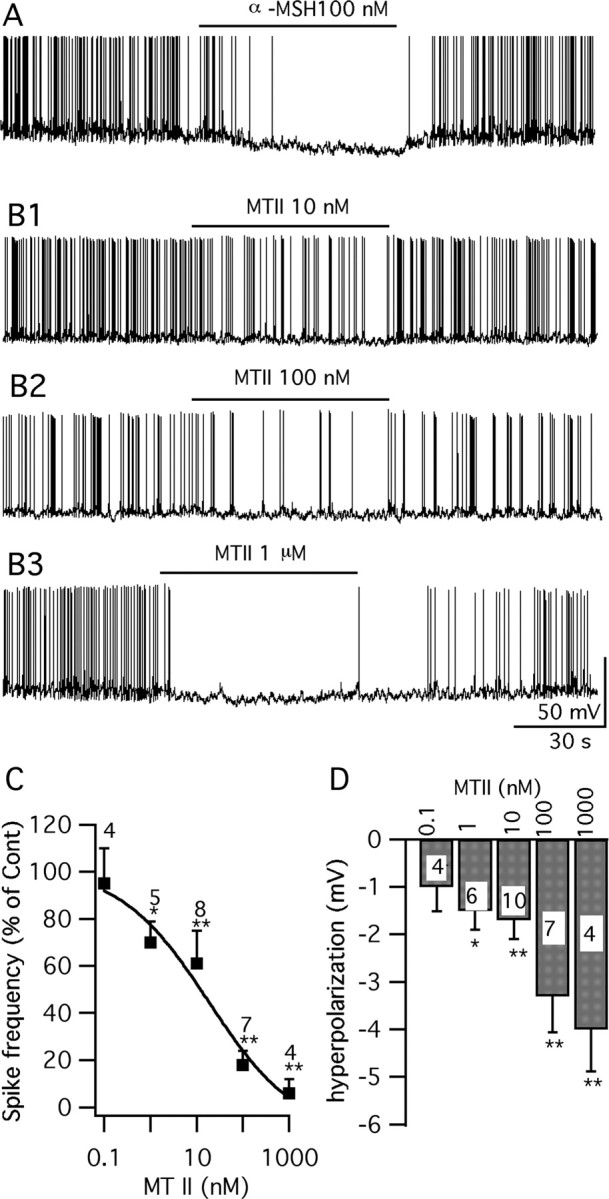
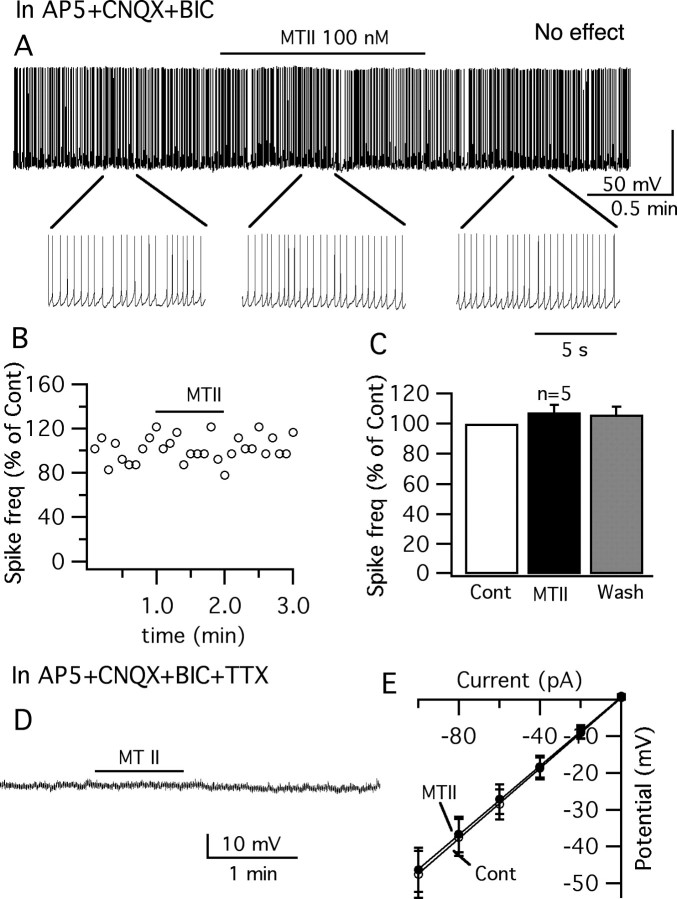
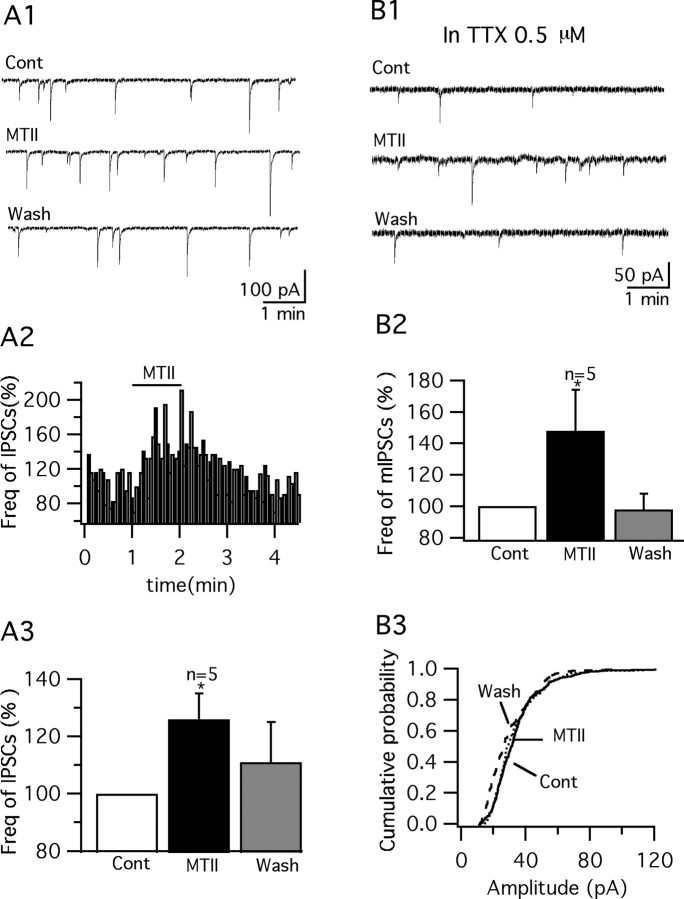
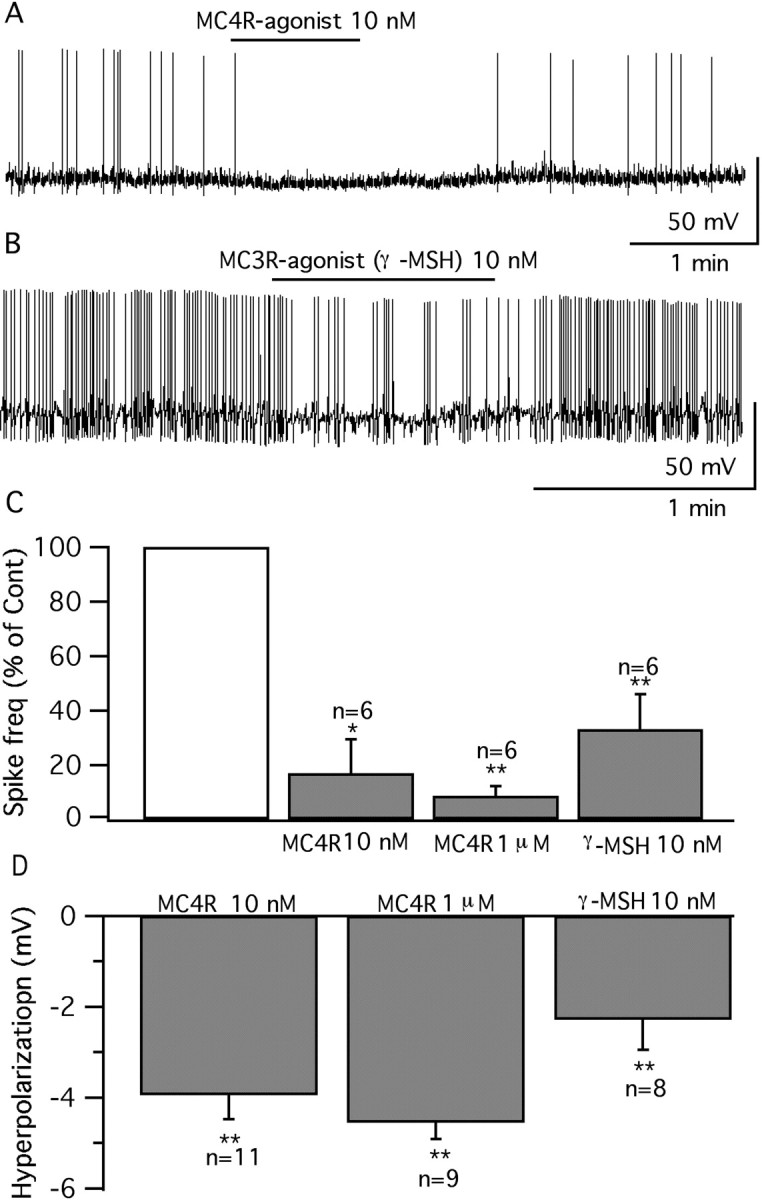
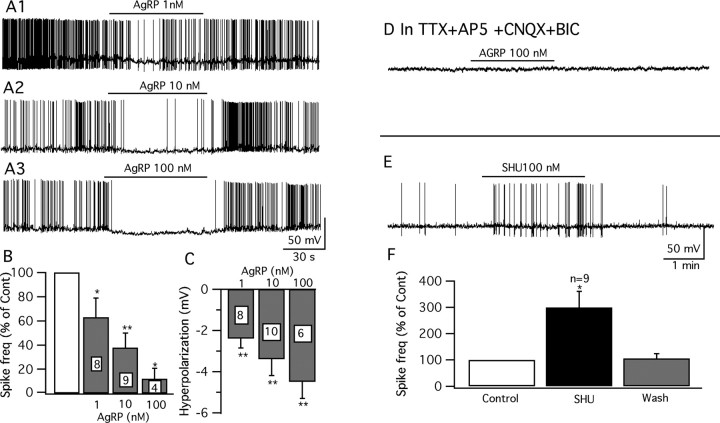
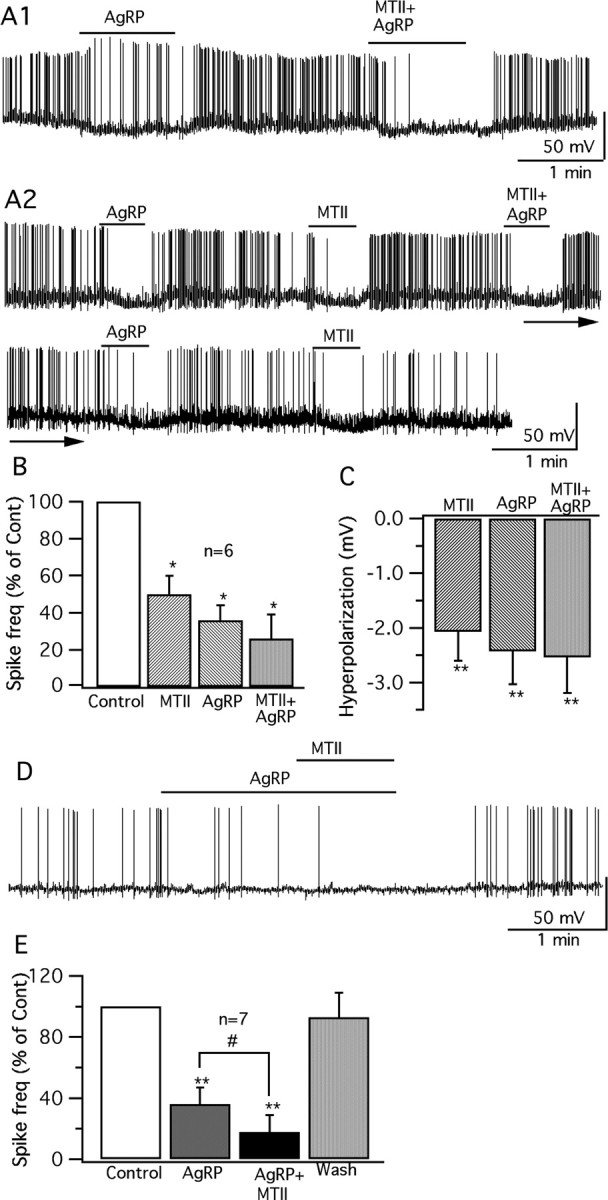
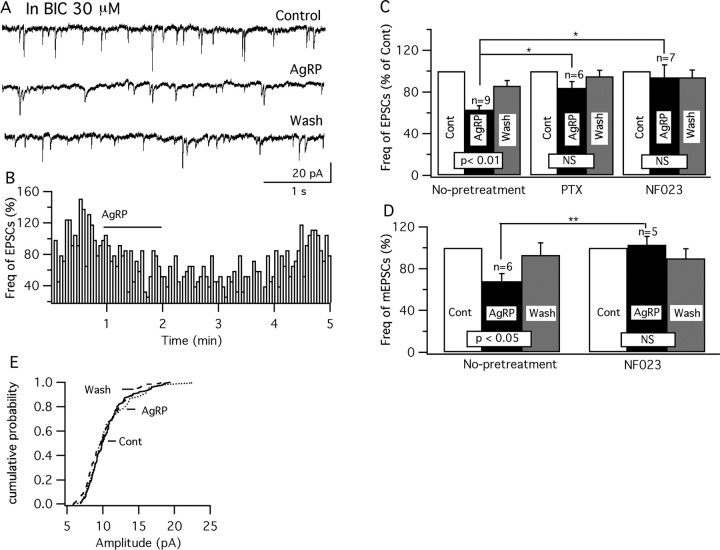
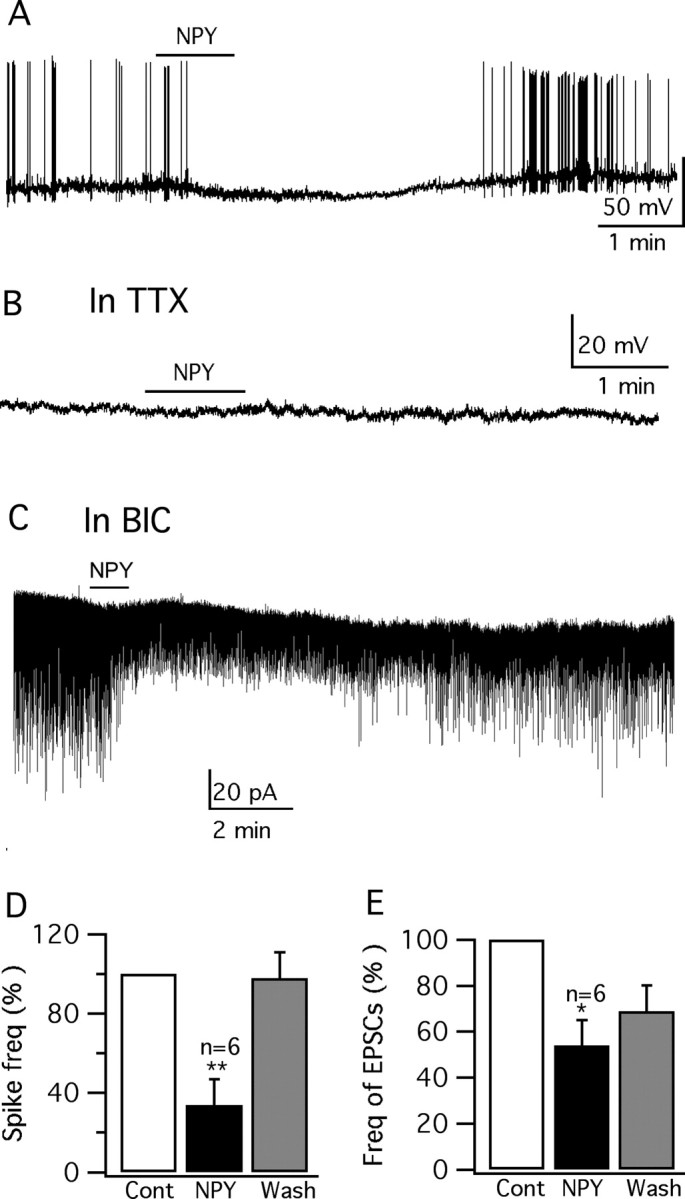
Anorexigenic melanocortins decrease food intake by activating MC3/MC4 receptors (MC3/4R); the prevailing view is that the orexigenic neuropeptide agouti-related peptide (AgRP) exerts the opposite action by acting as an antagonist at MC3/MC4 receptors. A total of 370 hypothalamic ventromedial nucleus (VMH) glutamatergic neurons was studied using whole-cell recording in hypothalamic slices from a novel mouse expressing green fluorescent protein (GFP) under control of the vesicular glutamate transporter 2 (vGluT2) promoter. Massive numbers of GFP-expressing VMH dendrites extended out of the core of the nucleus into the surrounding cell-poor shell. VMH dendrites received frequent appositions from AgRP-immunoreactive axons in the shell of the nucleus, but not the core, suggesting that AgRP may influence target VMH neurons. alpha-MSH, melanotan II (MTII), and selective MC3R or MC4R agonists were all inhibitory, reducing the spontaneous firing rate and hyperpolarizing vGluT2 neurons. The MC3/4R antagonist SHU9119 was excitatory. Unexpectedly, AgRP did not attenuate MTII actions on these neurons; instead, these two compounds showed an additive inhibitory effect. In the absence of synaptic activity, no hyperpolarization or change in input resistance was evoked by either MTII or AgRP, suggesting indirect actions. Consistent with this view, MTII increased the frequency of spontaneous and miniature IPSCs. In contrast, the mechanism of AgRP inhibition was dependent on presynaptic inhibition of EPSCs mediated by G(i)/G(o)-proteins, and was attenuated by pertussis toxin and NF023, inconsistent with mediation by G(s)-proteins associated with MC receptors. Together, our data suggest that the mechanism of AgRP actions on these excitatory VMH cells appears to be independent of the actions of melanocortins on MC receptors.
Figures
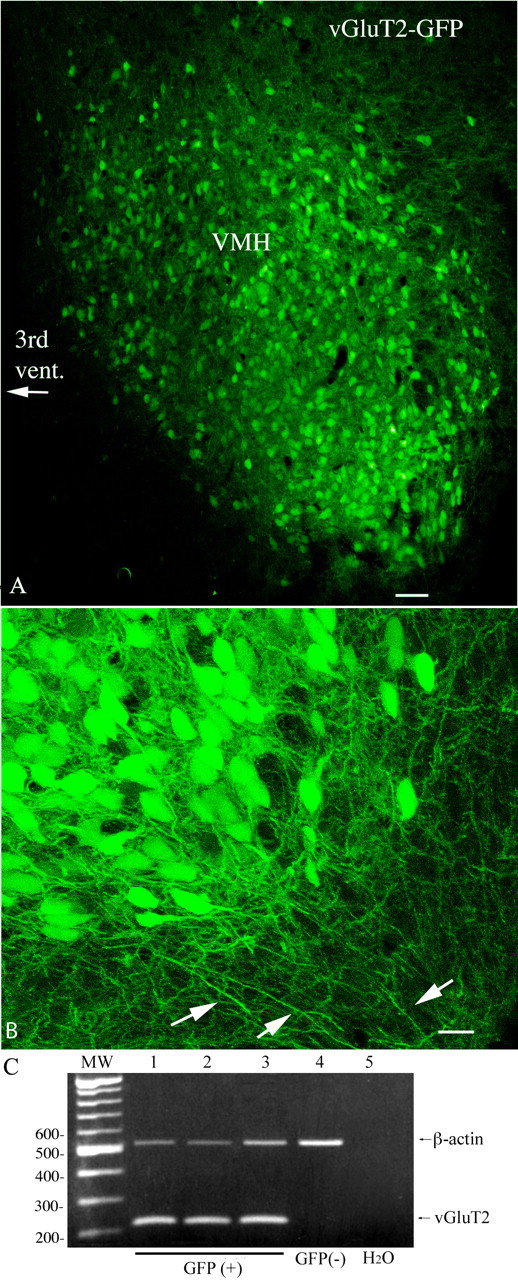
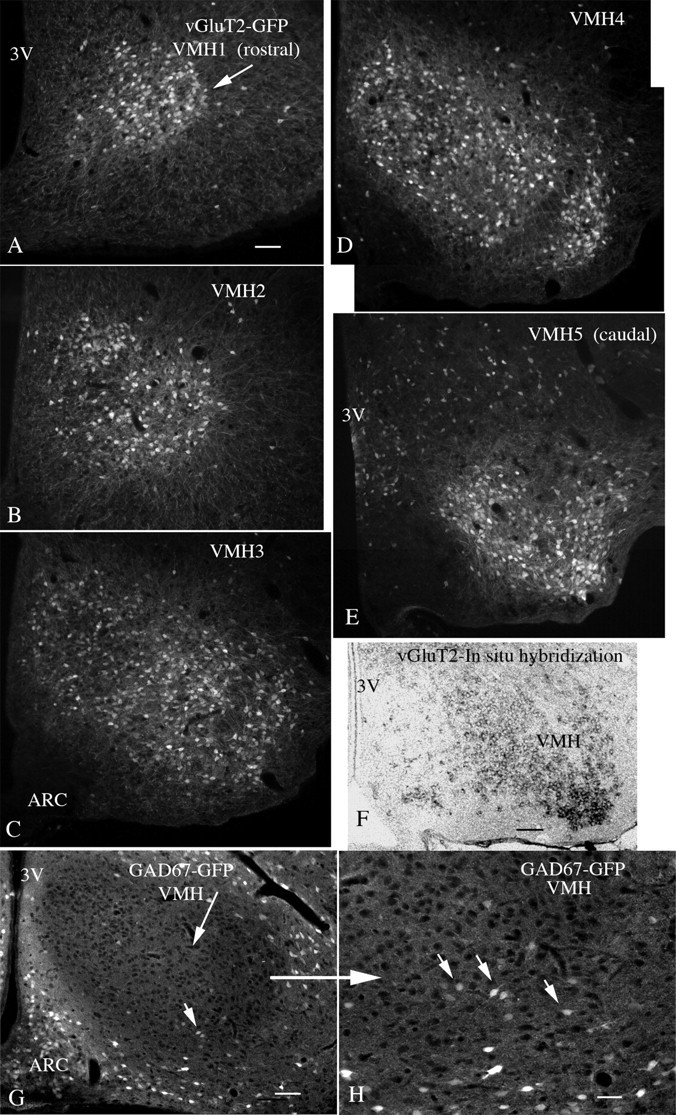
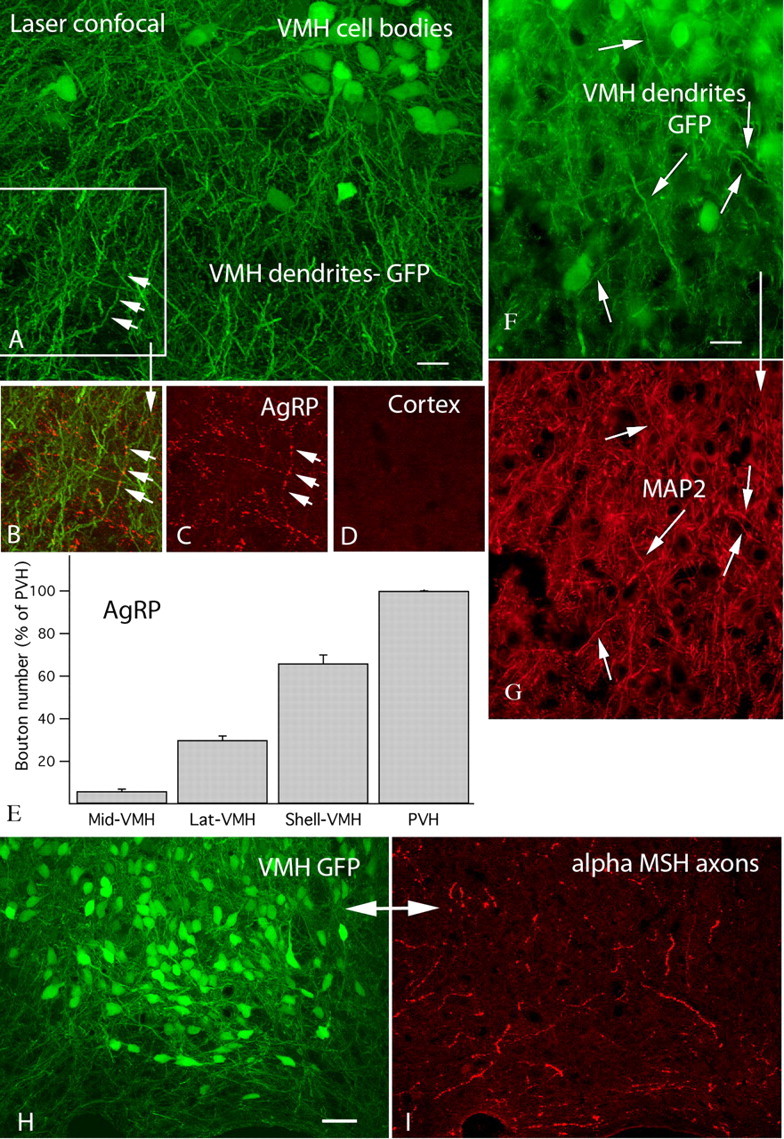
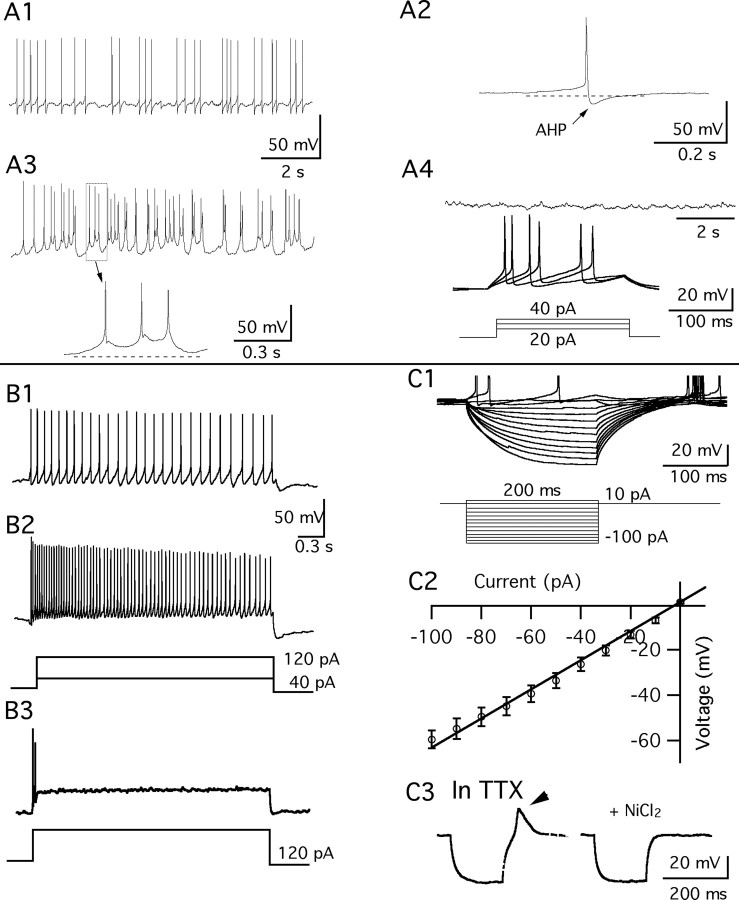
References
-
- Acuna-Goycolea C, Tamamaki N, Yanagawa Y, Obata K, van den Pol AN. Mechanisms of neuropeptide Y, peptide YY, and pancreatic polypeptide inhibition of identified green fluorescent protein-expressing GABA neurons in the hypothalamic neuroendocrine arcuate nucleus. J Neurosci. 2005;25:7406–7419. - PMC - PubMed
-
- Beart PM, Nicolopoulos LS, West DC, Headley PM. An excitatory amino acid projection from ventromedial hypothalamus to periaqueductal gray in the rat: autoradiographic and electrophysiological evidence. Neurosci Lett. 1988;85:205–211. - PubMed
-
- Bednarek MA, MacNeil T, Tang R, Kalyani RN, Van der Ploeg LH, Weinberg DH. Potent and selective peptide agonists of alpha-melanotropin action at human melanocortin receptor 4: their synthesis and biological evaluation in vitro. Biochem Biophys Res Commun. 2001;286:641–645. - PubMed
-
- Beindl W, Mitterauer T, Hohenegger M, Ijzerman AP, Nanoff C, Freissmuth M. Inhibition of receptor/G protein coupling by suramin analogues. Mol Pharmacol. 1996;50:415–423. - PubMed
-
- Bekkers JM, Stevens CF. Quantal analysis of EPSCs recorded from small numbers of synapses in hippocampal cultures. J Neurophysiol. 1995;73:1145–1156. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources