

Action-based body maps in the spinal cord emerge from a transitory floating organization
- PMID: 18495883
- PMCID: PMC6670632
- DOI: 10.1523/JNEUROSCI.0651-08.2008
Action-based body maps in the spinal cord emerge from a transitory floating organization
Abstract
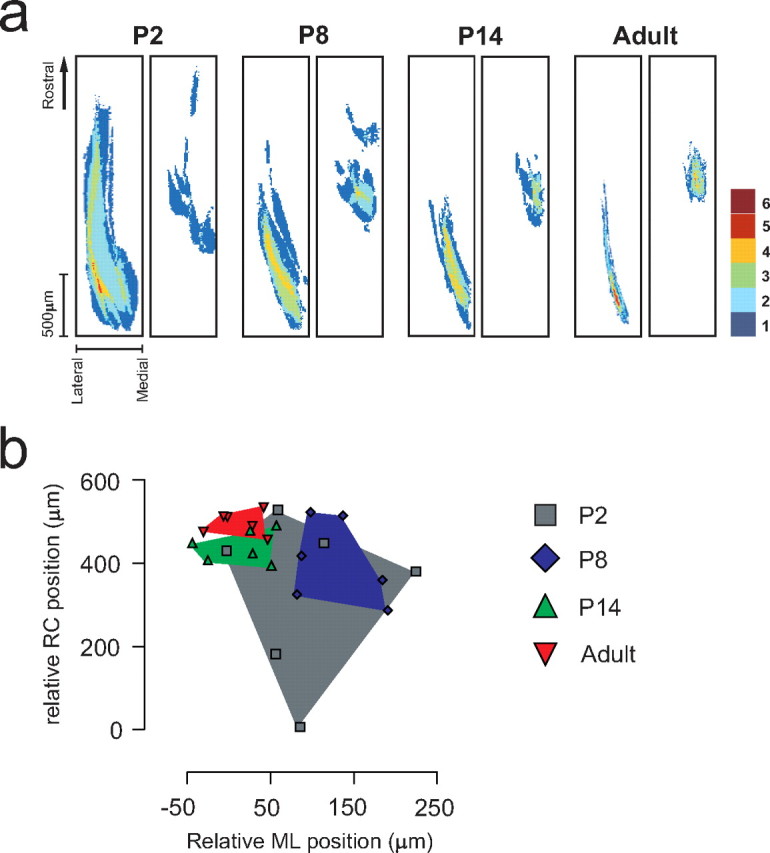
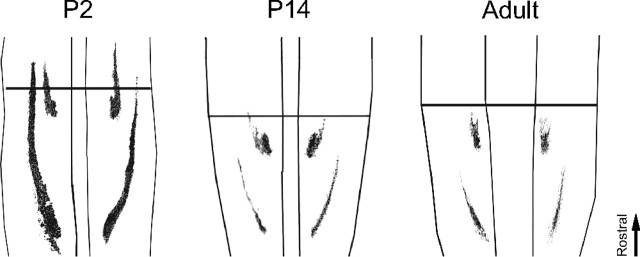
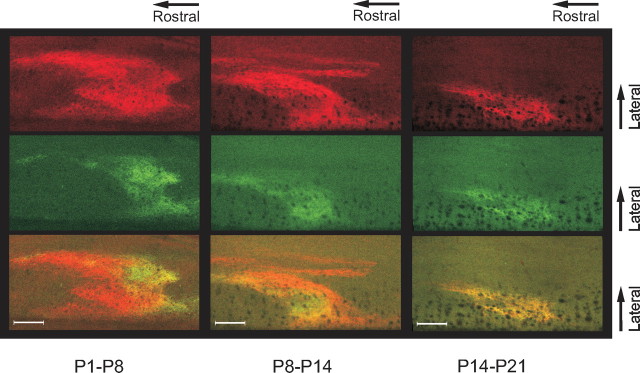
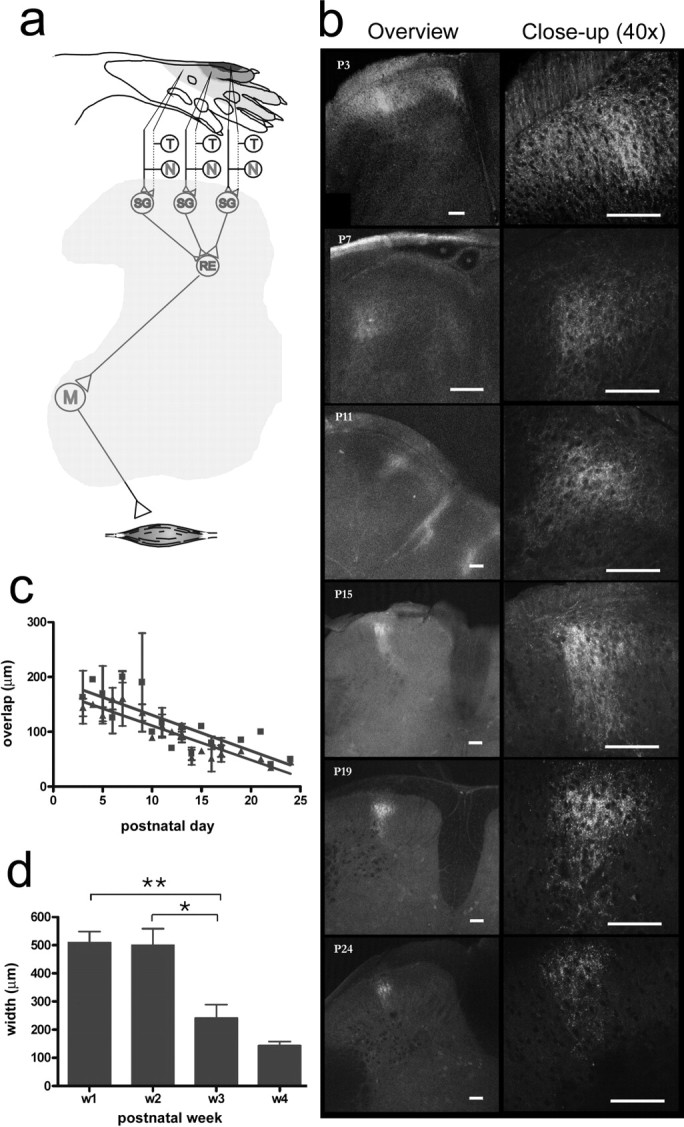
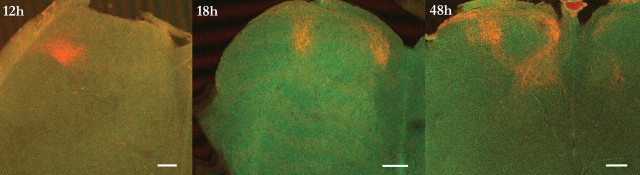
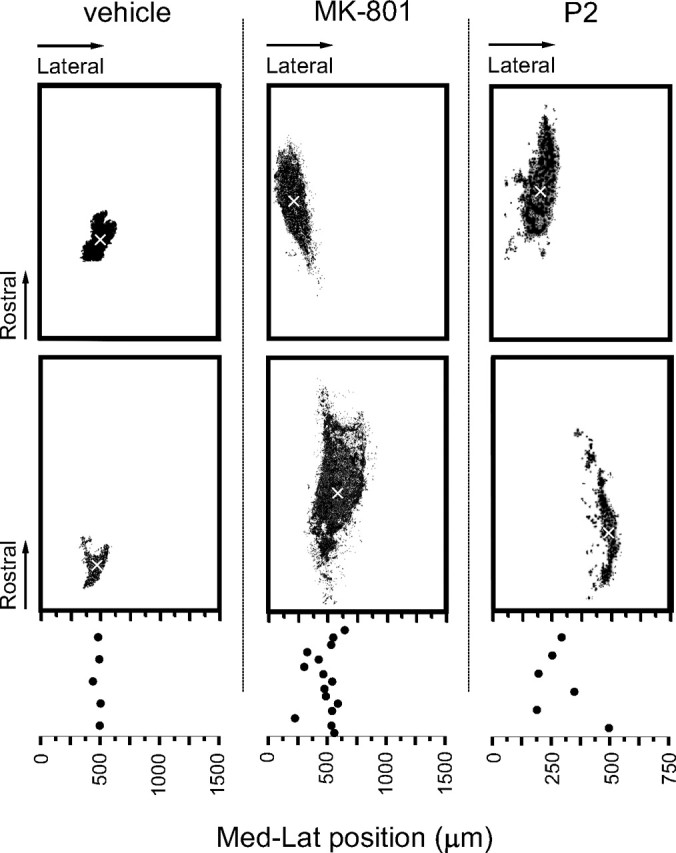
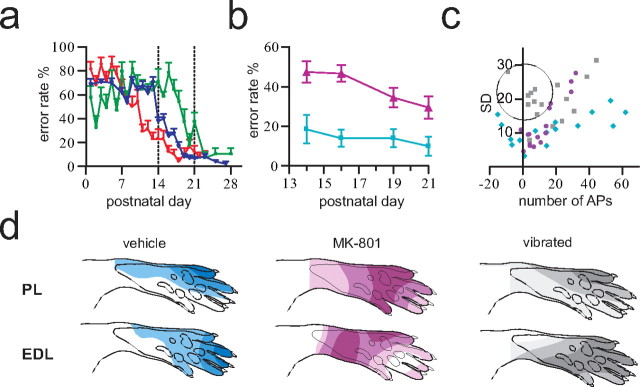
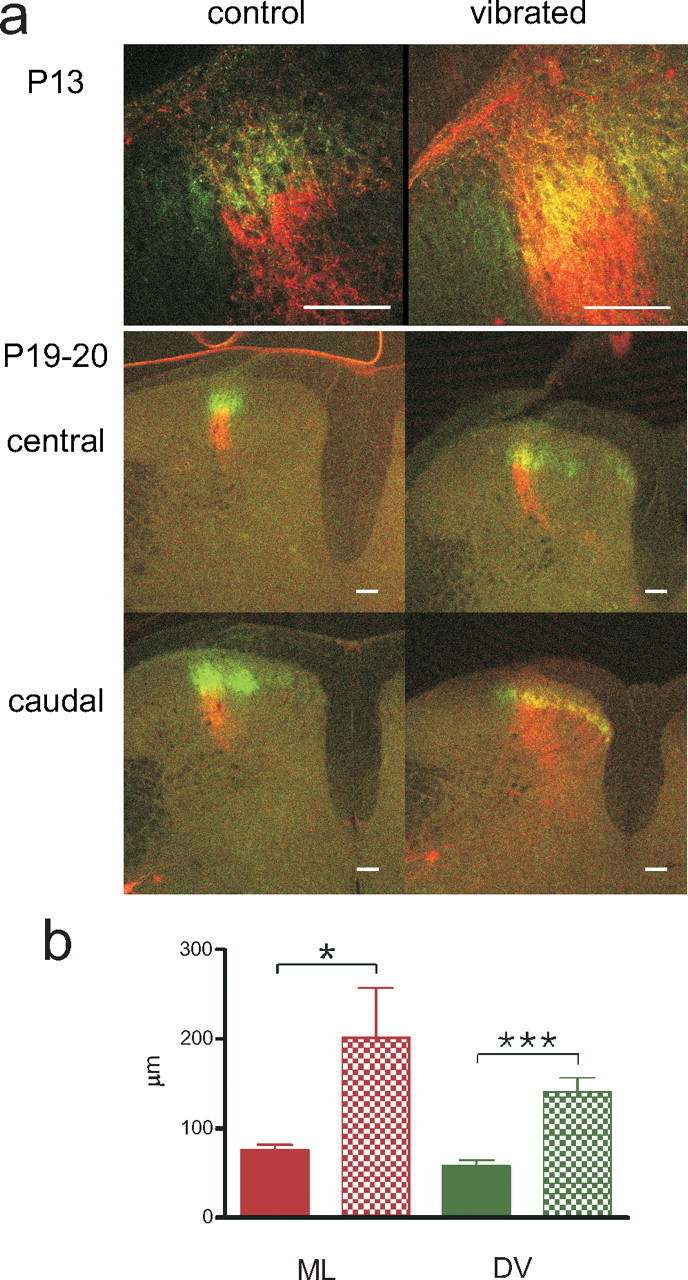
During development primary afferents grow into and establish neuronal connections in the spinal cord, thereby forming the basis for how we perceive sensory information and control our movements. In the somatosensory system, myriads of primary afferents, conveying information from different body locations and sensory modalities, get organized in the dorsal horn of the spinal cord so that spinal multisensory circuits receive topographically ordered information. How this intricate pathfinding is brought about during development is, however, largely unknown. Here we show that a body representation closely related to motor patterns emerges from a transitory floating and plastic organization through profound activity-dependent rewiring, involving both sprouting and elimination of afferent connections, and provide evidence for cross-modality interactions in the alignment of the multisensory input. Thus, far from being inborn and stereotypic, the dorsal horn of the spinal cord now appears to be a highly adaptive brain-body interface.
Figures
References
-
- Altman J, Bayer SA. The development of the rat spinal cord. Adv Anat Embryol Cell Biol. 1984;85:1–164. - PubMed
-
- Beggs S, Torsney C, Drew LJ, Fitzgerald M. The postnatal reorganization of primary afferent input and dorsal horn cell receptive fields in the rat spinal cord is an activity-dependent process. Eur J Neurosci. 2002;16:1249–1258. - PubMed
-
- Brown PB, Koerber HR, Millecchia R. Assembly of the dorsal horn somatotopic map. Somatosens Mot Res. 1997;14:93–106. - PubMed
-
- Brown PB, Koerber HR, Millecchia R. From innervation density to tactile acuity: 1. Spatial representation. Brain Res. 2004;1011:14–32. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources