

Prefrontal-inferotemporal interaction is not always necessary for reversal learning
- PMID: 18495887
- PMCID: PMC6670622
- DOI: 10.1523/JNEUROSCI.0952-08.2008
Prefrontal-inferotemporal interaction is not always necessary for reversal learning
Abstract
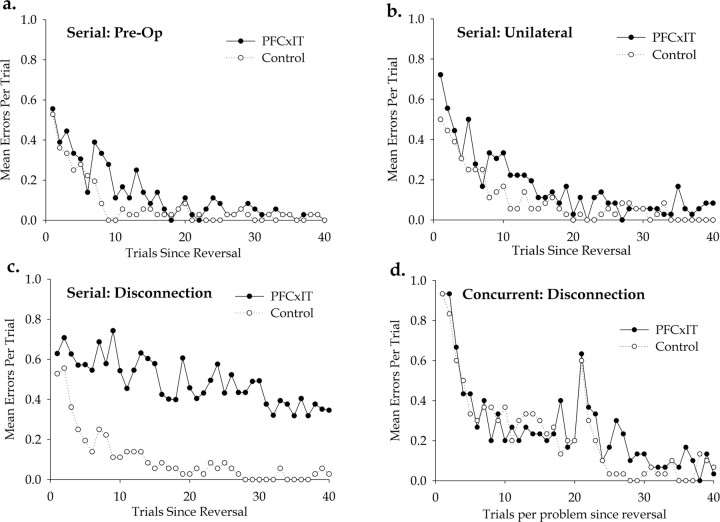
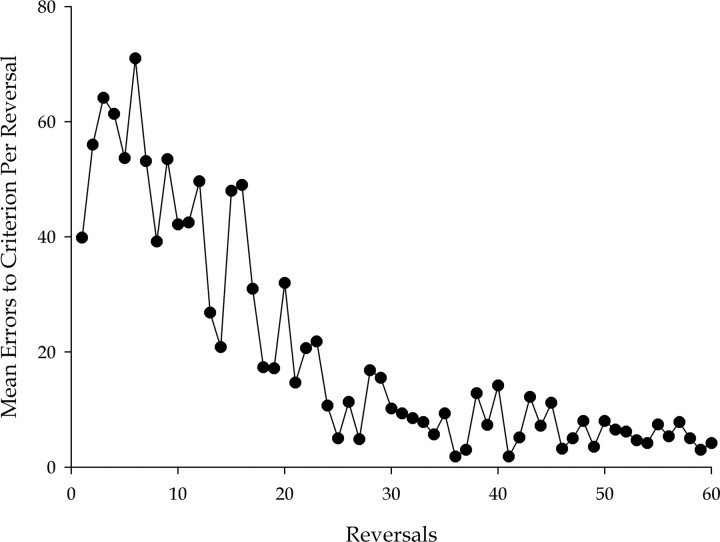
Prefrontal cortex (PFC) is thought to have a wide-ranging role in cognition, often described as executive function or behavioral inhibition. A specific example of such a role is the inhibition of representations in more posterior regions of cortex in a "top-down" manner, a function thought to be tested by reversal learning tasks. The direct action of PFC on posterior regions can be directly tested by disconnecting PFC from the region in question. We tested whether PFC directly inhibits visual object representations in inferotemporal cortex (IT) during reversal learning by studying the effect, in macaque monkeys, of disconnecting PFC from IT by crossed unilateral ablations. We tested two visual object reversal learning tasks, namely serial and concurrent reversal learning. We found that the disconnection severely impairs serial reversal learning but leaves concurrent reversal learning completely intact. Thus, PFC cannot be said to always have direct inhibitory control over visual object representations in reversal learning. Furthermore, our results cannot be explained by generalized theories of PFC function such as executive function and behavioral inhibition, because those theories do not make predictions that differentiate different forms of reversal learning. The results do, however, support our proposal, based on other experimental evidence from macaque monkeys, that PFC has a highly specific role in the representation of temporally complex events.
Conflict of interest statement
The authors declare no competing financial interests.
Figures
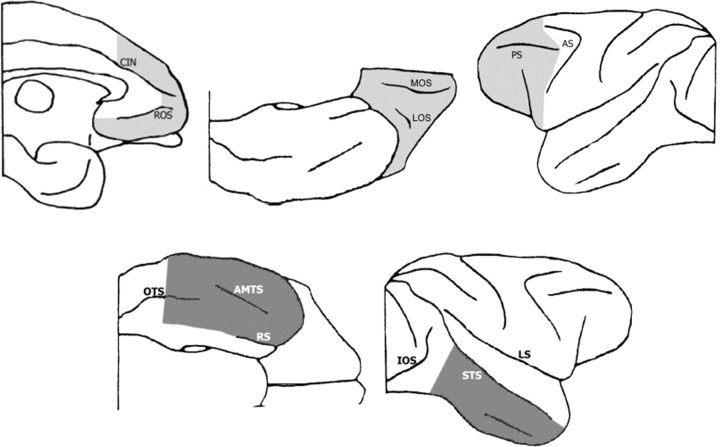
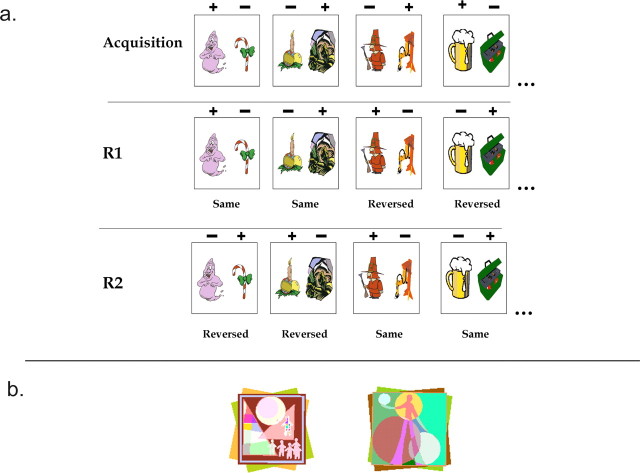
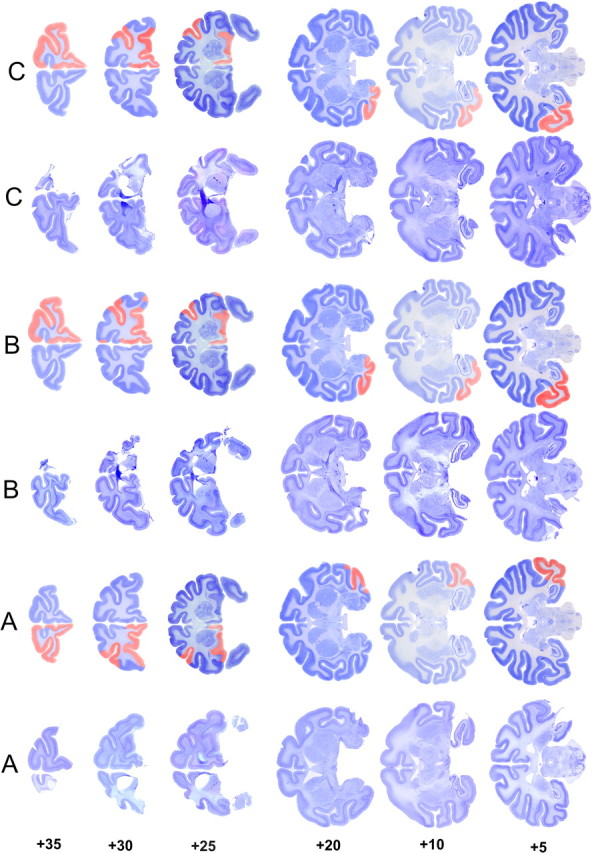
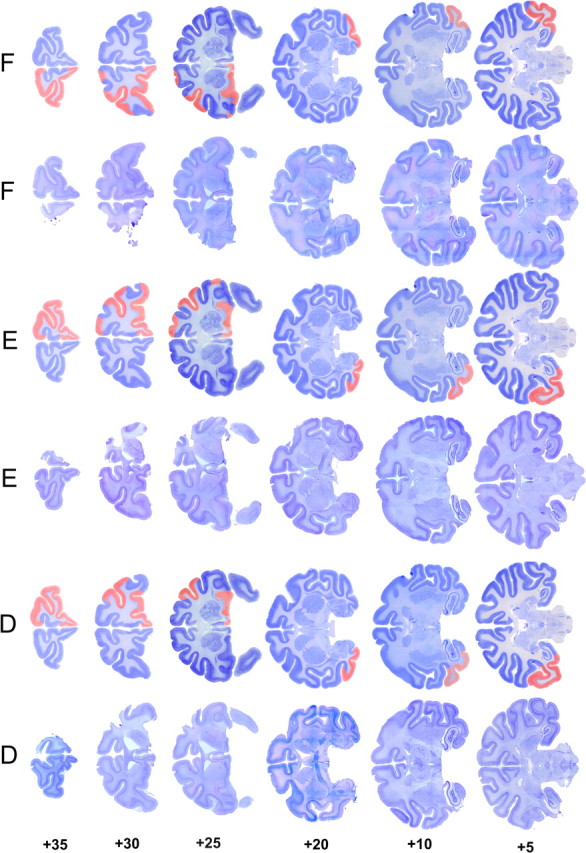
Similar articles
-
Prefrontal cortex function in the representation of temporally complex events.J Neurosci. 2008 Apr 9;28(15):3934-40. doi: 10.1523/JNEUROSCI.0633-08.2008. J Neurosci. 2008. PMID: 18400892 Free PMC article.
-
Reward-related reversal learning after surgical excisions in orbito-frontal or dorsolateral prefrontal cortex in humans.J Cogn Neurosci. 2004 Apr;16(3):463-78. doi: 10.1162/089892904322926791. J Cogn Neurosci. 2004. PMID: 15072681
-
Prefrontal-temporal disconnection impairs recognition memory but not familiarity discrimination.J Neurosci. 2013 Jun 5;33(23):9667-74. doi: 10.1523/JNEUROSCI.5759-12.2013. J Neurosci. 2013. PMID: 23739963 Free PMC article.
-
An analysis of rat prefrontal cortex in mediating executive function.Neurobiol Learn Mem. 2011 Oct;96(3):417-31. doi: 10.1016/j.nlm.2011.07.002. Epub 2011 Aug 9. Neurobiol Learn Mem. 2011. PMID: 21855643 Review.
-
Differential regulation of fronto-executive function by the monoamines and acetylcholine.Cereb Cortex. 2007 Sep;17 Suppl 1:i151-60. doi: 10.1093/cercor/bhm066. Cereb Cortex. 2007. PMID: 17725997 Review.
Cited by
-
Tract-Specific White Matter Correlates of Age-Related Reward Devaluation Deficits in Macaque Monkeys.J Neuroimaging Psychiatry Neurol. 2018;3(2):13-26. doi: 10.17756/jnpn.2018-023. Epub 2018 Jul 19. J Neuroimaging Psychiatry Neurol. 2018. PMID: 30198011 Free PMC article.
-
Ventral striatum lesions do not affect reinforcement learning with deterministic outcomes on slow time scales.Behav Neurosci. 2017 Oct;131(5):385-91. doi: 10.1037/bne0000211. Epub 2017 Aug 14. Behav Neurosci. 2017. PMID: 28805428 Free PMC article.
-
Orbitofrontal cortex volume in area 11/13 predicts reward devaluation, but not reversal learning performance, in young and aged monkeys.J Neurosci. 2014 Jul 23;34(30):9905-16. doi: 10.1523/JNEUROSCI.3918-13.2014. J Neurosci. 2014. PMID: 25057193 Free PMC article.
-
Ventrolateral prefrontal cortex is required for performance of a strategy implementation task but not reinforcer devaluation effects in rhesus monkeys.Eur J Neurosci. 2009 May;29(10):2049-59. doi: 10.1111/j.1460-9568.2009.06740.x. Epub 2009 May 9. Eur J Neurosci. 2009. PMID: 19453635 Free PMC article.
-
Behavioral flexibility is associated with changes in structure and function distributed across a frontal cortical network in macaques.PLoS Biol. 2020 May 26;18(5):e3000605. doi: 10.1371/journal.pbio.3000605. eCollection 2020 May. PLoS Biol. 2020. PMID: 32453728 Free PMC article.
References
-
- Aron AR, Robbins TW, Poldrack RA. Inhibition and the right inferior frontal cortex. Trends Cogn Sci. 2004;8:170–177. - PubMed
-
- Baxter MG, Browning PG. Two wrongs make a right: deficits in reversal learning after orbitofrontal damage are improved by amygdala ablation. Neuron. 2007;54:1–3. - PubMed
-
- Browning PG, Easton A, Buckley MJ, Gaffan D. The role of prefrontal cortex in object-in-place learning in monkeys. Eur J Neurosci. 2005;22:3281–3291. - PubMed
-
- Browning PGF, Easton A, Gaffan D. Frontal-temporal disconnection abolishes object discrimination learning set in macaque monkeys. Cereb Cortex. 2007;17:859–864. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous