

The iron-sulphur protein Ind1 is required for effective complex I assembly
- PMID: 18497740
- PMCID: PMC2435133
- DOI: 10.1038/emboj.2008.98
The iron-sulphur protein Ind1 is required for effective complex I assembly
Abstract
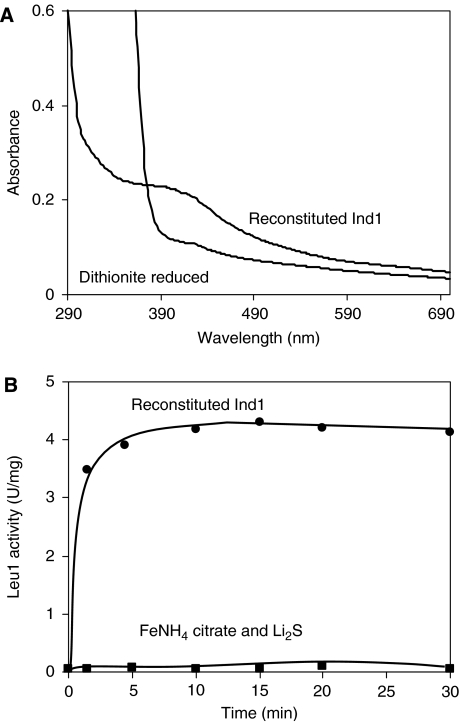
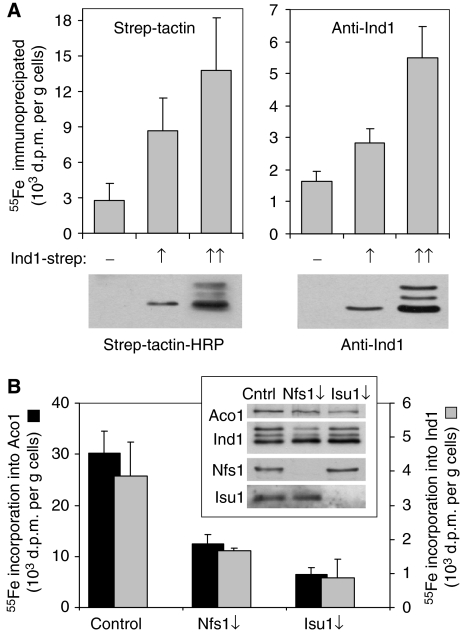
NADH:ubiquinone oxidoreductase (complex I) of the mitochondrial inner membrane is a multi-subunit protein complex containing eight iron-sulphur (Fe-S) clusters. Little is known about the assembly of complex I and its Fe-S clusters. Here, we report the identification of a mitochondrial protein with a nucleotide-binding domain, named Ind1, that is required specifically for the effective assembly of complex I. Deletion of the IND1 open reading frame in the yeast Yarrowia lipolytica carrying an internal alternative NADH dehydrogenase resulted in slower growth and strongly decreased complex I activity, whereas the activities of other mitochondrial Fe-S enzymes, including aconitase and succinate dehydrogenase, were not affected. Two-dimensional gel electrophoresis, in vitro activity tests and electron paramagnetic resonance signals of Fe-S clusters showed that only a minor fraction (approximately 20%) of complex I was assembled in the ind1 deletion mutant. Using in vivo and in vitro approaches, we found that Ind1 can bind a [4Fe-4S] cluster that was readily transferred to an acceptor Fe-S protein. Our data suggest that Ind1 facilitates the assembly of Fe-S cofactors and subunits of complex I.
Figures
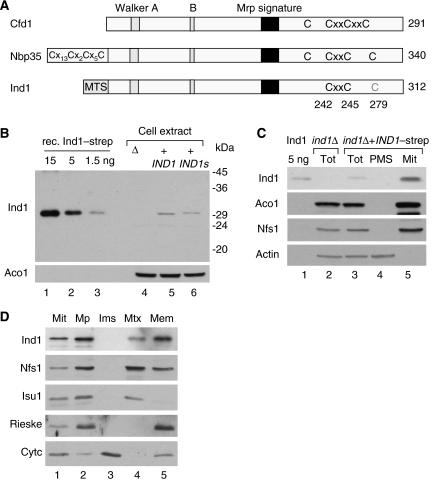
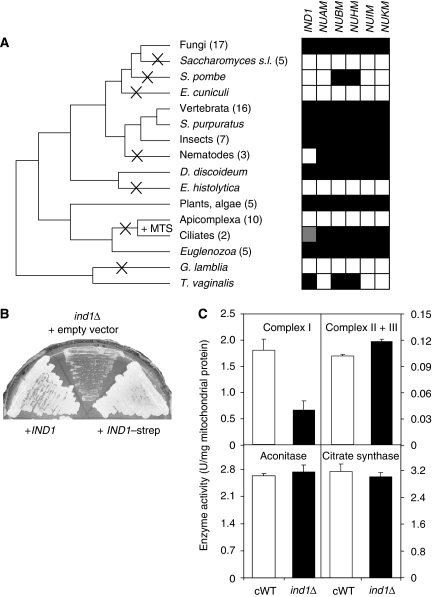
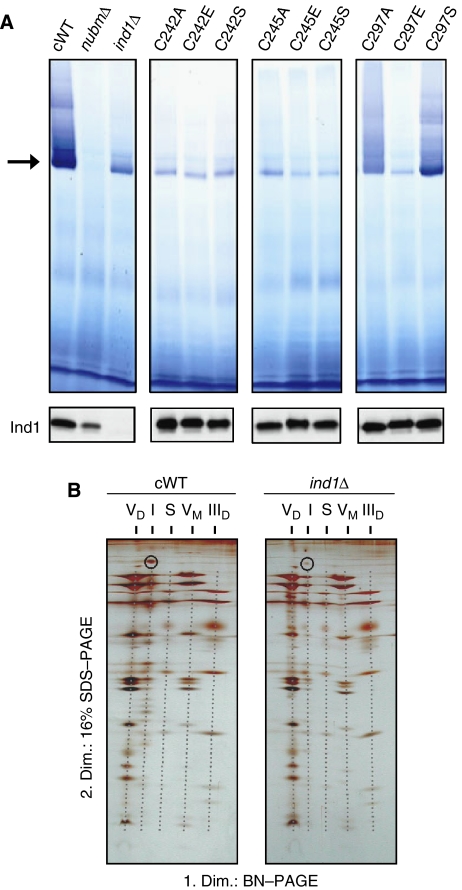
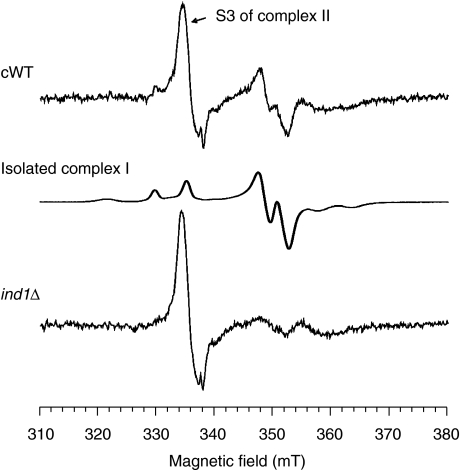
References
-
- Abdrakhmanova A, Dobrynin K, Zwicker K, Kerscher S, Brandt U (2006) Functional sulfurtransferase is associated with mitochondrial complex I from Yarrowia lipolytica, but is not required for assembly of its iron–sulfur clusters. FEBS Lett 579: 6781–6785 - PubMed
-
- Balk J, Lobréaux S (2005) Biogenesis of iron–sulfur proteins in plants. Trends Plant Sci 10: 324–331 - PubMed
-
- Beinert H (2000) Iron–sulfur proteins: ancient structures, still full of surprises. J Biol Inorg Chem 5: 2–15 - PubMed
-
- Brandt U (2006) Energy converting NADH:quinone oxidoreductase (complex I). Annu Rev Biochem 75: 69–92 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
