

ATM promotes the obligate XY crossover and both crossover control and chromosome axis integrity on autosomes
- PMID: 18497861
- PMCID: PMC2374915
- DOI: 10.1371/journal.pgen.1000076
ATM promotes the obligate XY crossover and both crossover control and chromosome axis integrity on autosomes
Abstract
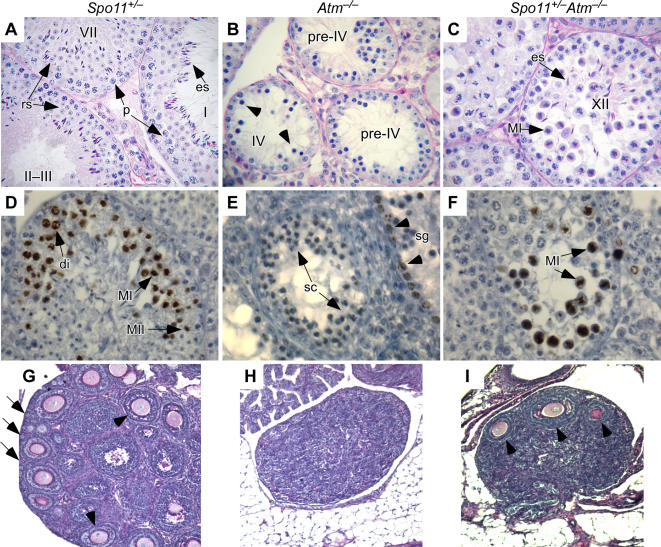
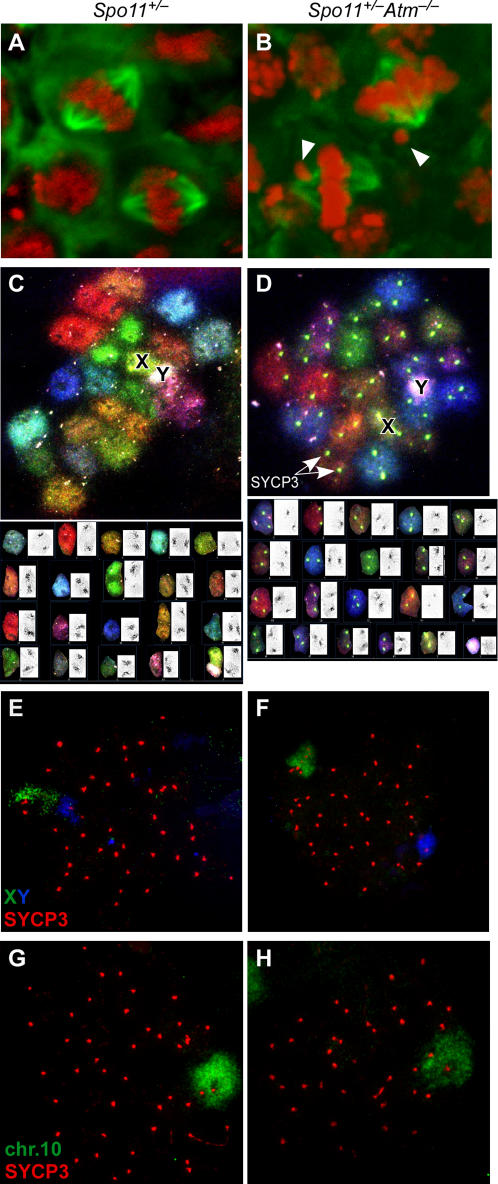
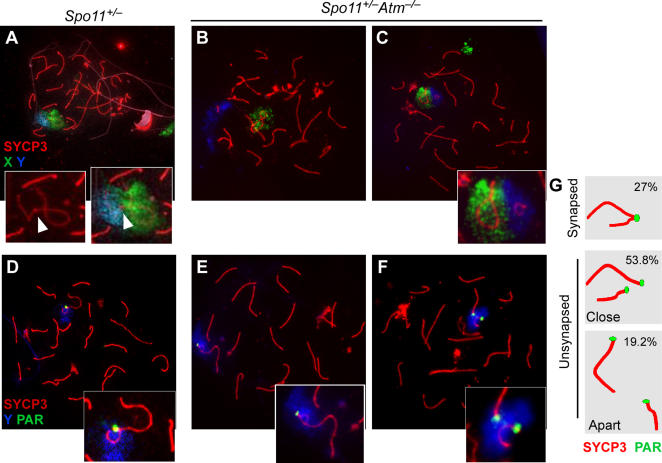
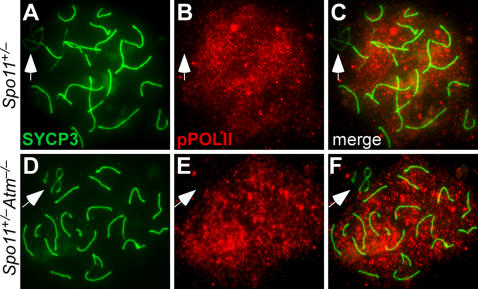
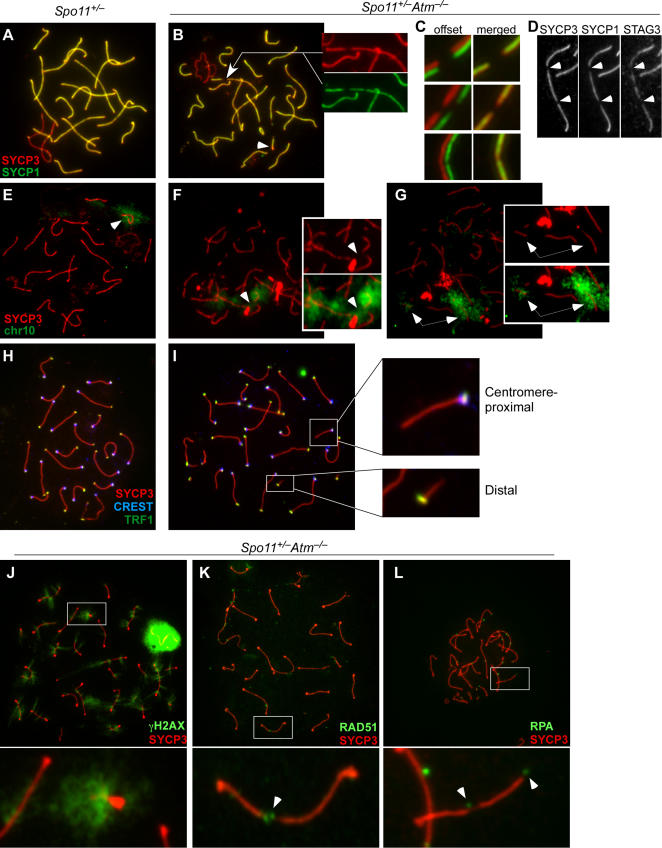
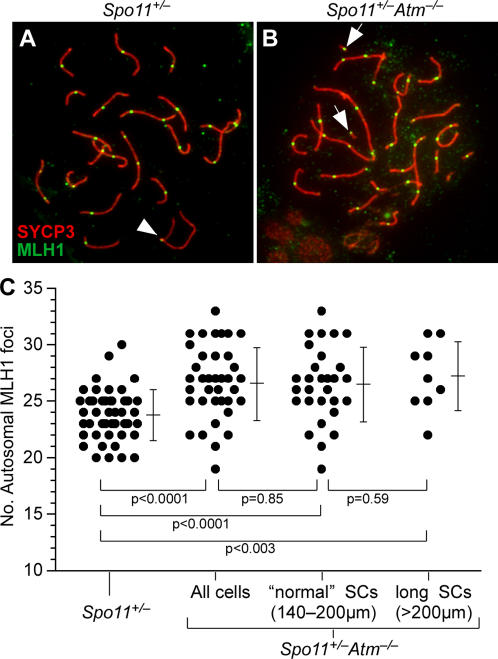
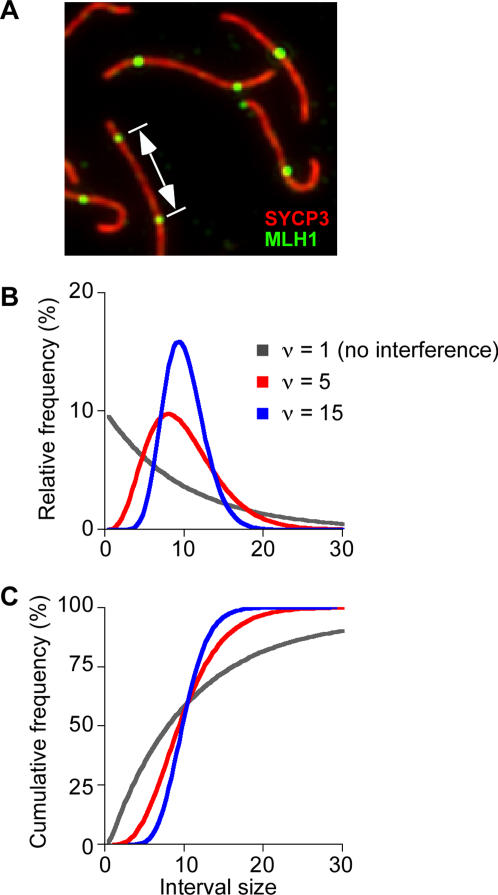
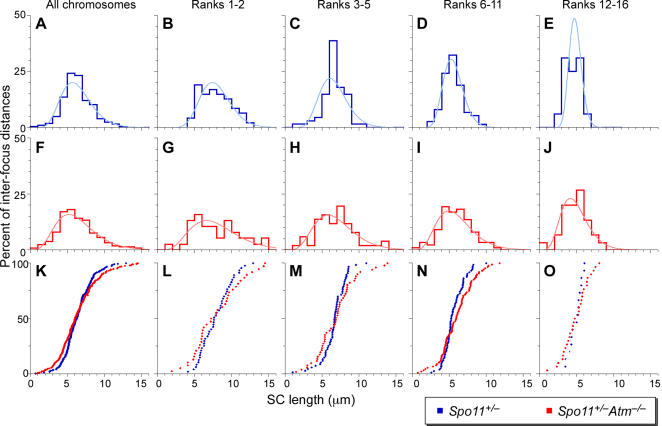
During meiosis in most sexually reproducing organisms, recombination forms crossovers between homologous maternal and paternal chromosomes and thereby promotes proper chromosome segregation at the first meiotic division. The number and distribution of crossovers are tightly controlled, but the factors that contribute to this control are poorly understood in most organisms, including mammals. Here we provide evidence that the ATM kinase or protein is essential for proper crossover formation in mouse spermatocytes. ATM deficiency causes multiple phenotypes in humans and mice, including gonadal atrophy. Mouse Atm-/- spermatocytes undergo apoptosis at mid-prophase of meiosis I, but Atm(-/-) meiotic phenotypes are partially rescued by Spo11 heterozygosity, such that ATM-deficient spermatocytes progress to meiotic metaphase I. Strikingly, Spo11+/-Atm-/- spermatocytes are defective in forming the obligate crossover on the sex chromosomes, even though the XY pair is usually incorporated in a sex body and is transcriptionally inactivated as in normal spermatocytes. The XY crossover defect correlates with the appearance of lagging chromosomes at metaphase I, which may trigger the extensive metaphase apoptosis that is observed in these cells. In addition, control of the number and distribution of crossovers on autosomes appears to be defective in the absence of ATM because there is an increase in the total number of MLH1 foci, which mark the sites of eventual crossover formation, and because interference between MLH1 foci is perturbed. The axes of autosomes exhibit structural defects that correlate with the positions of ongoing recombination. Together, these findings indicate that ATM plays a role in both crossover control and chromosome axis integrity and further suggests that ATM is important for coordinating these features of meiotic chromosome dynamics.
Conflict of interest statement
The authors have declared that no competing interests exist.
Figures
References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
