

Glutamate receptor dynamics in dendritic microdomains
- PMID: 18498731
- PMCID: PMC2572138
- DOI: 10.1016/j.neuron.2008.04.030
Glutamate receptor dynamics in dendritic microdomains
Abstract
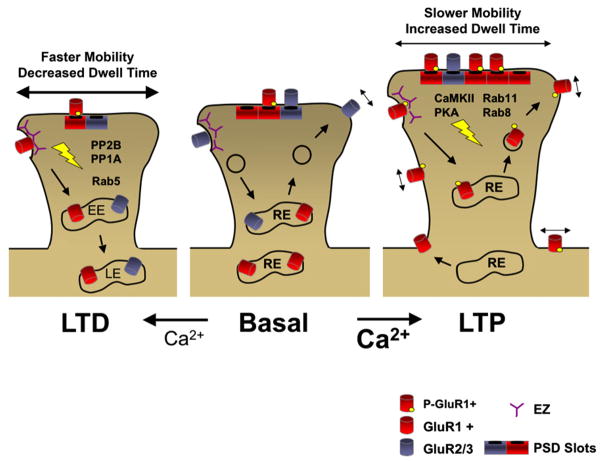
Among diverse factors regulating excitatory synaptic transmission, the abundance of postsynaptic glutamate receptors figures prominently in molecular memory and learning-related synaptic plasticity. To allow for both long-term maintenance of synaptic transmission and acute changes in synaptic strength, the relative rates of glutamate receptor insertion and removal must be tightly regulated. Interactions with scaffolding proteins control the targeting and signaling properties of glutamate receptors within the postsynaptic membrane. In addition, extrasynaptic receptor populations control the equilibrium of receptor exchange at synapses and activate distinct signaling pathways involved in plasticity. Here, we review recent findings that have shaped our current understanding of receptor mobility between synaptic and extrasynaptic compartments at glutamatergic synapses, focusing on AMPA and NMDA receptors. We also examine the cooperative relationship between intracellular trafficking and surface diffusion of glutamate receptors that underlies the expression of learning-related synaptic plasticity.
Figures
References
-
- Adesnik H, Nicoll RA, England PM. Photoinactivation of native AMPA receptors reveals their real-time trafficking. Neuron. 2005;48:977–985. - PubMed
-
- Akaaboune M, Culican SM, Turney SG, Lichtman JW. Rapid and reversible effects of activity on acetylcholine receptor density at the neuromuscular junction in vivo. Science. 1999;286:503–507. - PubMed
-
- Akaaboune M, Grady RM, Turney S, Sanes JR, Lichtman JW. Neurotransmitter receptor dynamics studied in vivo by reversible photounbinding of fluorescent ligands. Neuron. 2002;34:865–876. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
