

Cells on the run: shear-regulated integrin activation in leukocyte rolling and arrest on endothelial cells
- PMID: 18499427
- PMCID: PMC5912339
- DOI: 10.1016/j.ceb.2008.04.003
Cells on the run: shear-regulated integrin activation in leukocyte rolling and arrest on endothelial cells
Abstract
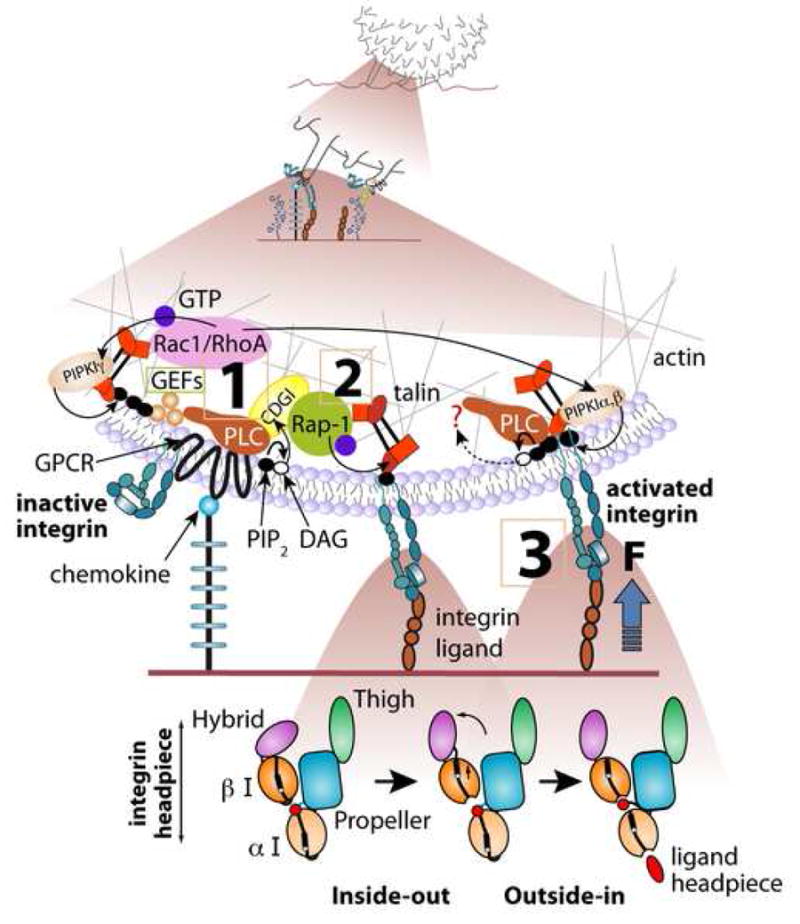
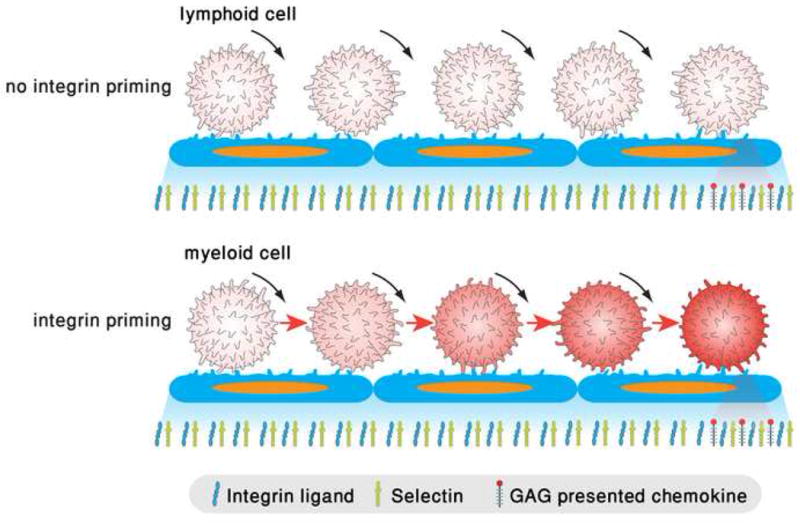
The arrest of rolling leukocytes on various target vascular beds is mediated by specialized leukocyte integrins and their endothelial immunoglobulin superfamily (IgSF) ligands. These integrins are kept in largely inactive states and undergo in situ activation upon leukocyte-endothelial contact by both biochemical and mechanical signals from flow-derived shear forces. In vivo and in vitro studies suggest that leukocyte integrin activation involves conformational alterations through inside-out signaling followed by ligand-induced rearrangements accelerated by external forces. This activation process takes place within fractions of seconds by in situ signals transduced to the rolling leukocyte as it encounters specialized endothelial-displayed chemoattractants, collectively termed arrest chemokines. In neutrophils, selectin rolling engagements trigger intermediate affinity integrins to support reversible adhesions before chemokine-triggered arrest. Different leukocyte subsets appear to use different modalities of integrin activation during rolling and arrest at distinct endothelial sites.
Figures
References
-
- Springer TA. Traffic signals for lymphocyte recirculation and leukocyte emigration: The multistep paradigm. Cell. 1994;76:301–314. - PubMed
-
- Constantin G, Majeed M, Giagulli C, Piccio L, Kim JY, Butcher EC, Laudanna C. Chemokines trigger immediate β2 integrin affinity and mobility changes: differential regulation and roles in lymphocyte arrest under flow. Immunity. 2000;13:759–769. - PubMed
-
- Rot A, von Andrian UH. Chemokines in innate and adaptive host defense: basic chemokinese grammar for immune cells. Annu Rev Immunol. 2004;22:891–928. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
