

Triple-helix structure in telomerase RNA contributes to catalysis
- PMID: 18500353
- PMCID: PMC2562722
- DOI: 10.1038/nsmb.1420
Triple-helix structure in telomerase RNA contributes to catalysis
Abstract
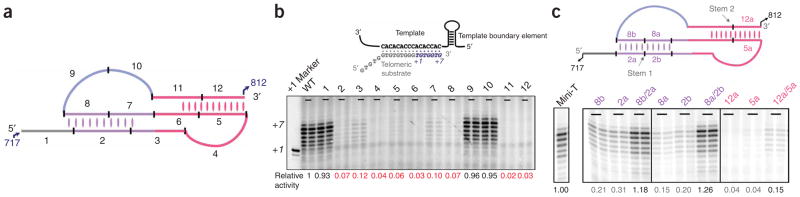
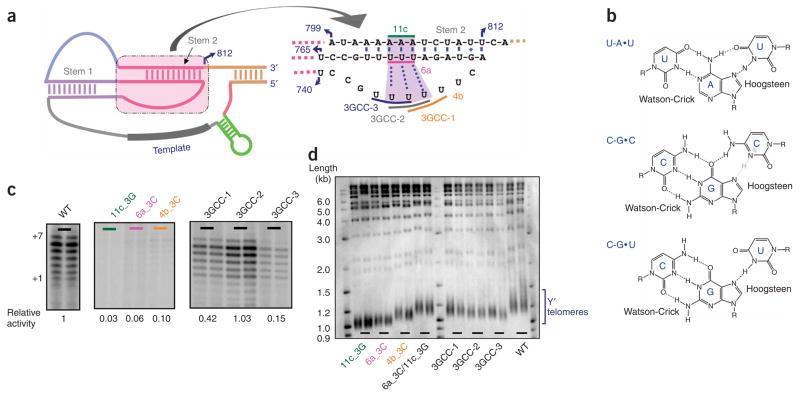
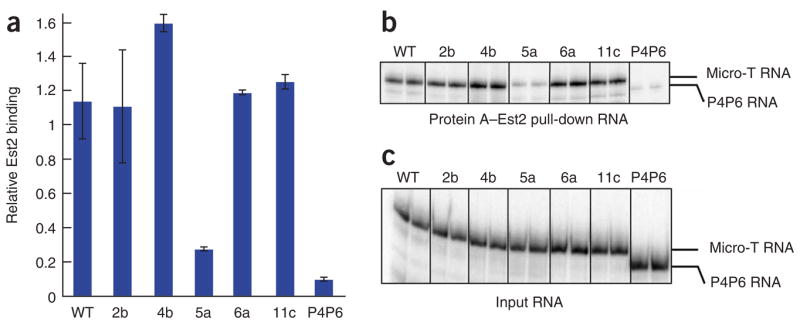
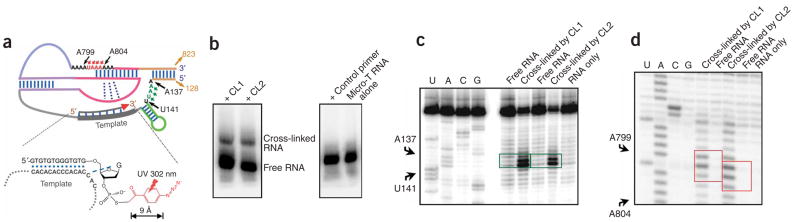
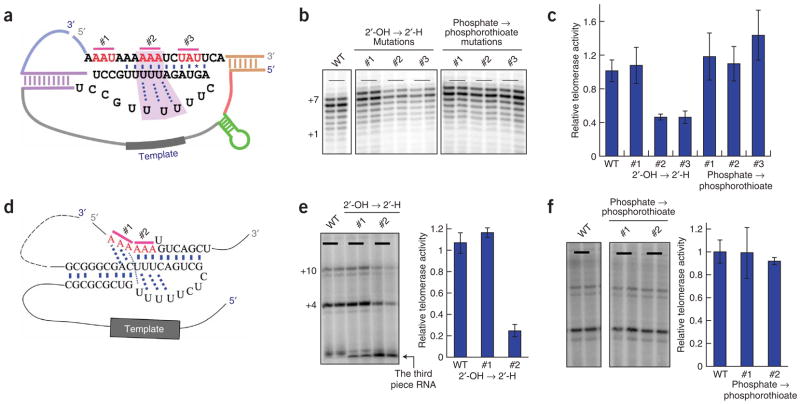
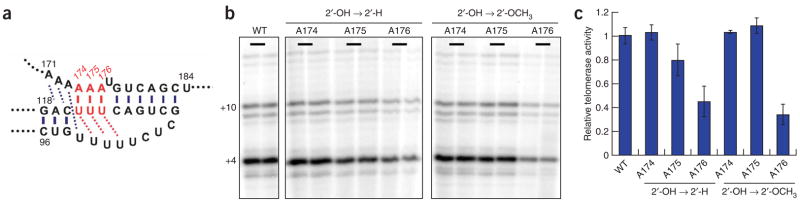
Telomerase is responsible for replication of the ends of linear chromosomes in most eukaryotes. Its intrinsic RNA subunit provides the template for synthesis of telomeric DNA by the reverse-transcriptase (TERT) subunit and tethers other proteins into the ribonucleoprotein (RNP) complex. We report that a phylogenetically conserved triple helix within a pseudoknot structure of this RNA contributes to telomerase activity but not by binding the TERT protein. Instead, 2'-OH groups protruding from the triple helix participate in both yeast and human telomerase catalysis; they may orient the primer-template relative to the active site in a manner analogous to group I ribozymes. The role of RNA in telomerase catalysis may have been acquired relatively recently or, alternatively, telomerase may be a molecular fossil representing an evolutionary link between RNA enzymes and RNP enzymes.
Figures
References
-
- Autexier C, Lue NF. The structure and function of telomerase reverse transcriptase. Annu Rev Biochem. 2006;75:493–517. - PubMed
-
- Cech TR. Beginning to understand the end of the chromosome. Cell. 2004;116:273–279. - PubMed
-
- Singer MS, Gottschling DE. TLC1: template RNA component of Saccharomyces cerevisiae telomerase. Science. 1994;266:404–409. - PubMed
-
- Lingner J, et al. Reverse transcriptase motifs in the catalytic subunit of telomerase. Science. 1997;276:561–567. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
