

doi: 10.1128/EC.00132-08.
Epub 2008 May 23.
Pet191 is a cytochrome c oxidase assembly factor in Saccharomyces cerevisiae
Affiliations
- PMID: 18503002
- PMCID: PMC2519776
- DOI: 10.1128/EC.00132-08
Item in Clipboard
Pet191 is a cytochrome c oxidase assembly factor in Saccharomyces cerevisiae
Eukaryot Cell.
2008 Aug.
Abstract
The twin-Cx(9)C motif protein Pet191 is essential for cytochrome c oxidase maturation. The motif Cys residues are functionally important and appear to be present in disulfide linkages within a large oligomeric complex associated with the mitochondrial inner membrane. The import of Pet191 differs from that of other twin-Cx(9)C motif class of proteins in being independent of the Mia40 pathway.
Figures
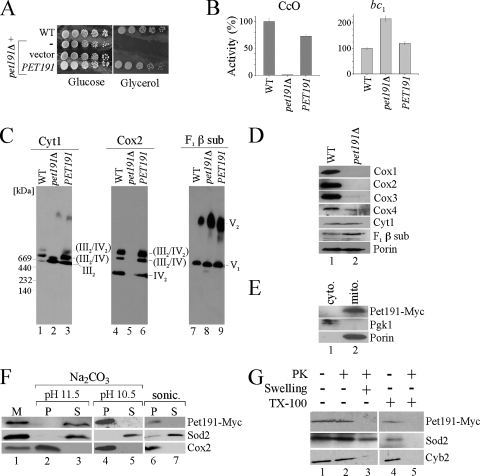
Pet191 is required for CcO biogenesis. (A) Serial dilutions of pet191Δ cells transformed with an empty vector or PET191 were plated onto complete medium containing 2% glucose or 2% lactate/glycerol as a carbon source. The WT and pet191Δ diploid strains of Saccharomyces cerevisiae (BY4743) were used. (B) CcO and SDH/bc1 activities. Activities of the WT mitochondria were set as 100%. Mitochondria were isolated from cells cultured on raffinose medium. (C) Mitochondria were solubilized in a lysis buffer containing 1.5% digitonin and protein complexes were separated by BN-PAGE. The respiratory chain complexes (III, IV, and V) were analyzed by immunoblotting with respective antibodies. sub, subunit. (D) Steady-state levels of mitochondrial proteins were analyzed by sodium dodecyl sulfate (SDS)-PAGE followed by Western blotting. (E) Cells expressing Pet191-Myc were fractionated, and cytosolic (cyto.) and mitochondrial (mito.) fractions were collected, resolved by SDS-PAGE, and analyzed by immunoblotting with the anti-Myc, anti-Pgk1, and anti-porin antibodies. The Pet191-Myc was expressed on a pRS426 vector under the MET25 promoter and the CYC1 terminator (17). (F) Mitochondria of a strain expressing Pet191-Myc (M; lane 1) were treated with either 0.1 M Na2CO3 (pH 10.5 or 11.5) or sonicated (sonic.) and fractionated by high-speed centrifugation. Obtained pellet (P; lanes 2, 4, and 6) and supernatant (S; lanes 3, 5, and 7) fractions were analyzed by immunoblotting using antibodies against the Myc epitope, matrix protein Sod2, and IM-anchored protein Cox2. (G) Intact (lanes 1, 2, and 4) or swollen (lane 3) mitochondria were incubated with (lanes 2, 3, and 5) or without (lanes 1 and 4) proteinase K (PK). In lanes 4 and 5, mitochondria were lysed with 1% Triton X-100 (TX-100) prior to incubation with PK. Western blot analysis was performed using antibodies specific for the Myc tag, matrix protein Sod2, and IMS protein Cyb2.

Molecular size of Pet191. (A) Mitochondria isolated from WT cells and the strain containing genomically tagged Pet191 (carrying PET191::3HA) were solubilized and subjected to BN-PAGE followed by immunoblot analysis with an antibody against the HA epitope. A genomically HA-tagged variant of PET191 was generated by homologous recombination, inserting the triple HA tag 3′ to the open reading frame. (B) Mitochondria prepared from WT and cox11Δ cells carrying PET191::3HA were analyzed by BN-PAGE as described previously (25) using anti-HA and porin antibodies. (C) Cells expressing Pet191-Myc or its mutant form (C5A) were used for mitochondrial preparation. Isolated mitochondria were lysed in a buffer containing 0.1% DOC in the presence (+) or absence (−) of 100 mM DTT, and clarified lysates were loaded onto the size exclusion column (10/30 G-200; Superdex) equilibrated with buffer containing 0.05% DOC ± 5 mM DTT. Fractions were directly assayed by slot immunoblotting with anti-Myc antibodies. Column void (VE) and internal (VI) volumes are marked by arrows.

Conserved cysteine residues are important for Pet191 function. (A) Serial dilutions of pet191Δ cells transformed with an empty vector, either PET191 or one of its mutant forms, were plated onto complete medium containing 2% glucose or 2% lactate/glycerol as a carbon source and grown for 4 days at 30°C or 37°C. Mutant forms of Pet191 (the C5A, C15A, C21A, C32A, C46A, C56A, and C86A mutants) were generated by site-directed mutagenesis using pRS426-PET191-Myc as a template. (B). Schematic demonstrating possible conformation of the Pet191 molecule where two Cx9C motifs face each other. Positions of the most critical cysteine residues are depicted by light gray circles. Other Cys residues are shown in black. (C) Mitochondria isolated from pet191Δ cells carrying an empty vector, PET191, or its mutant forms were subjected to SDS-PAGE followed by Western blot analysis with the antibodies against the Myc epitope and porin. (D) WT cells overexpressing PET191 or its C5A, C15A, or C56A mutant versions were serially diluted, plated as described for panel A, and grown for 3 days at 30°C. (E) Mitochondria from the cells coexpressing Pet191-HA and Pet191-Myc or its C5A mutant form were solubilized and analyzed as described above, except that DTT treatment was not included. Collected fractions were analyzed using anti-Myc and anti-HA antibodies. (F) Mitochondria isolated from the WT PET191::3HA strain with (lanes 1 to 3) or without (lanes 4 to 6) Pet191-Myc were lysed, and clarified extracts were immunoprecipitated with agarose-coupled anti-Myc antibodies. Co-IP of Pet191-Myc and Pet191-HA was performed as described previously (20), using anti-Myc-agarose-coupled beads (Santa Cruz Biotechnology). The load (representing 5% of the extracts; lanes 1 and 4), entire-wash (lanes 2 and 5), and eluate (lanes 3 and 6) fractions were analyzed by immunoblotting.
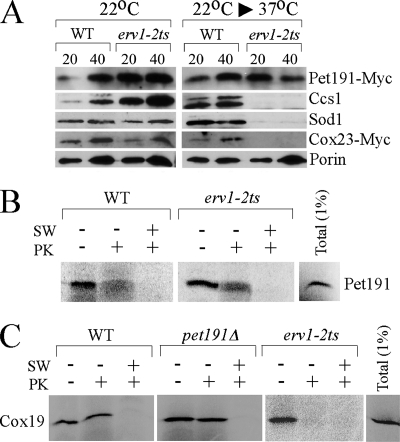
erv1 mutant mitochondria have normal levels of Pet191-Myc. (A) WT and temperature-sensitive erv1-2ts strains expressing Pet191-Myc or Cox23-Myc were grown at 22°C. The temperature-sensitive strain erv1-2ts (13) and the corresponding WT strain were a kind gift from Roland Lill. For isolation of mitochondria from erv1-2ts cells, the cells were pregrown in supplemented synthetic medium containing 2% raffinose at 22°C until the early logarithmic phase. Cultures were split into two halves, one of which was subjected to a temperature shift (22°C to 37°C), while the other was left at 22°C. After 7 to 10 h of incubation, cells were harvested and mitochondria isolated as described above. Mitochondria were isolated, separated by SDS-PAGE, and analyzed by immunoblotting with antibodies recognizing Myc-tag, Ccs1, Sod1, and porin. (B) Pet191 was synthesized in rabbit reticulocyte lysate in the presence of [35S]methionine and incubated for 20 min at 30°C with mitochondria isolated from either WT or erv1-2ts cells. After incubation, mitochondria were reisolated, washed, and either swollen in hypotonic medium (SW) or incubated with proteinase K (PK). Samples were then subjected to SDS-PAGE followed by autoradiography. (C) Radiolabeled Cox19 was imported into the WT, pet191Δ, and erv1-2ts mitochondria as described above.
References
-
- Abajian, C., L. A. Yatsunyk, B. E. Ramirez, and A. C. Rosenzweig. 2004. Yeast Cox17 solution structure and copper(I) binding. J. Biol. Chem. 27953584-53592. - PubMed
-
- Arnesano, F., E. Balatri, L. Banci, I. Bertini, and D. R. Winge. 2005. Folding studies of Cox17 reveal an important interplay of cysteine oxidase and copper binding. Structure 13713-722. - PubMed
-
- Barrientos, A., M. H. Barros, I. Valnot, A. Rotig, P. Rustin, and A. Tzagoloff. 2002. Cytochrome oxidase in health and disease. Gene 28653-63. - PubMed
-
- Barros, M. H., A. Johnson, and A. Tzagoloff. 2004. Cox23, a homologue of COX17, is required for cytochrome oxidase assembly. J. Biol. Chem. 27931943-31947. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
