

Insulin resistance in experimental alcohol-induced liver disease
- PMID: 18505416
- PMCID: PMC10012390
- DOI: 10.1111/j.1440-1746.2008.05339.x
Insulin resistance in experimental alcohol-induced liver disease
Abstract
Background and aim: Chronic ethanol consumption impairs liver regeneration due, in part, to inhibition of insulin signaling. This study characterizes the mechanisms and consequences of ethanol-impaired insulin signaling in relation to oxidative injury and altered gene expression.
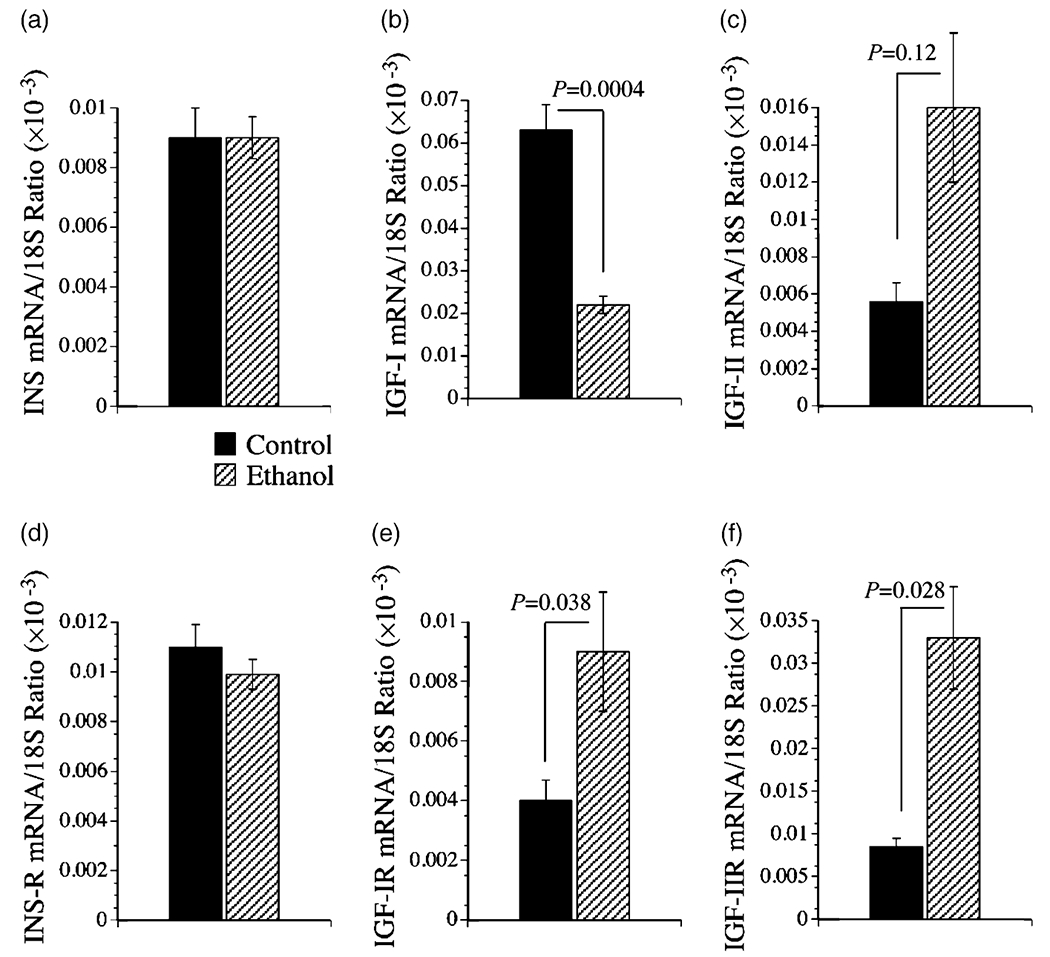
Methods: Long-Evans rats were fed for 8 weeks with isocaloric liquid diets containing 0% (control) or 37% ethanol (caloric content). Livers were used to examine histopathology, indices of oxidative stress, gene expression required for insulin and insulin-like growth factor (IGF) signaling, insulin-responsive gene expression, i.e. glyceraldehydes-3-phosphate dehydrogenase (GAPDH) and aspartyl-asparaginyl-beta-hydroxylase (AAH), and competitive equilibrium binding to the insulin, IGF-I, and IGF-II receptors.
Results: Chronic ethanol exposure caused liver injury with increased hepatocellular steatosis, inflammation, apoptosis, and increased immunoreactivity for activated caspase-3, 8-hydroxy-2'-deoxyguanosine, and 4-hydroxy-2,3-nonenol. These effects were associated with increased expression of IGF-I receptor, IGF-II, and IGF-II receptor, and expression of IGF-I, AAH, and GAPDH, which mediate energy metabolism and cell motility/remodeling, and reduced binding to the insulin receptor.
Conclusions: Chronic ethanol-induced liver injury causes insulin resistance with inhibition of insulin-responsive genes needed for metabolism, remodeling, and regeneration. In contrast, the IGF-I and IGF-II signaling mechanisms remain relatively preserved, suggesting that insulin-regulated hepatic functions may be selectively vulnerable to the toxic effects of ethanol.
Figures





References
-
- Wands JR, Carter EA, Bucher NL, Isselbacher KJ. Inhibition of hepatic regeneration in rats by acute and chronic ethanol intoxication. Gastroenterology 1979; 77: 528–31. - PubMed
-
- Diehl AM, Thorgeirsson SS, Steer CJ. Ethanol inhibits liver regeneration in rats without reducing transcripts of key protooncogenes. Gastroenterology 1990; 99: 1105–12. - PubMed
-
- Wands JR, Carter EA, Bucher NL, Isselbacher KJ. Effect of acute and chronic ethanol intoxication on hepatic regeneration. Adv. Exp. Med. Biol 1980; 132: 663–70. - PubMed
-
- Carter EA, Wands JR. Ethanol inhibits hormone stimulated hepatocyte DNA synthesis. Biochem. Biophys. Res. Commun 1985; 128: 767–74. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
