

Hypercholesterolemia impairs immunity to tuberculosis
- PMID: 18505807
- PMCID: PMC2493195
- DOI: 10.1128/IAI.00037-08
Hypercholesterolemia impairs immunity to tuberculosis
Abstract
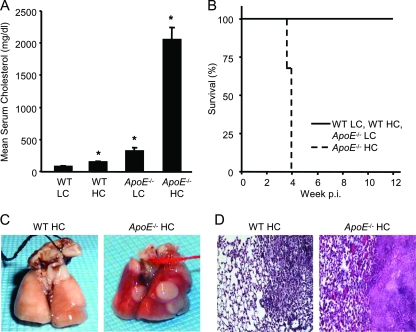
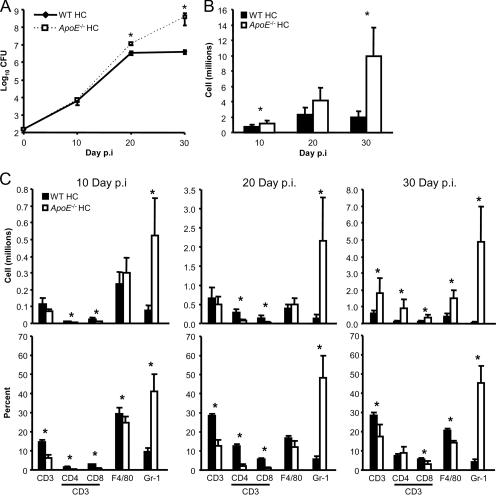
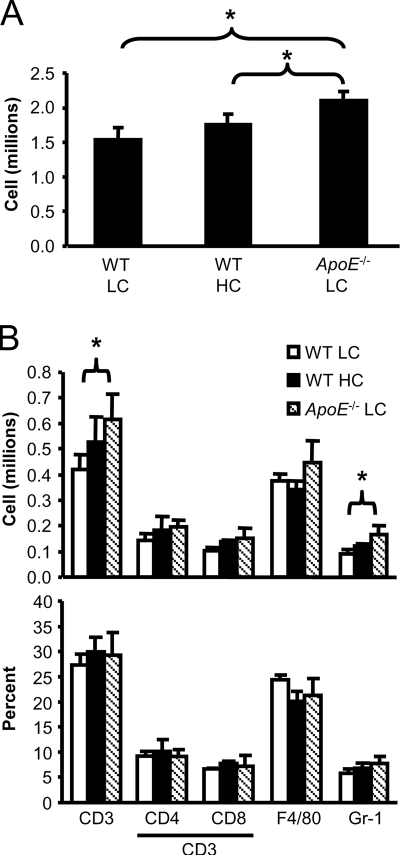
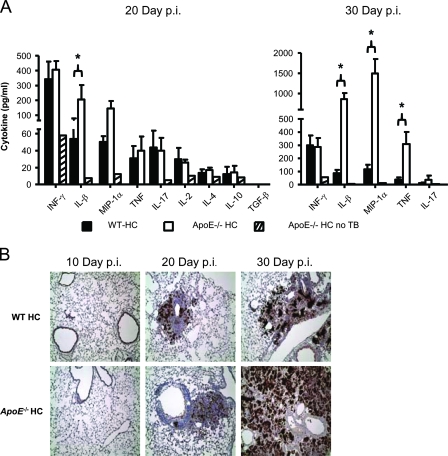
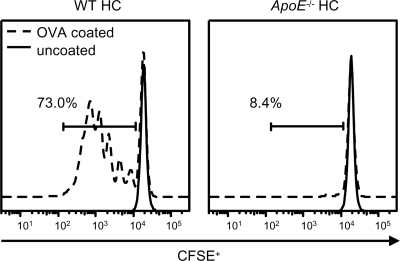
We demonstrate that apolipoprotein E -deficient (ApoE(-/-)) mice are highly susceptible to tuberculosis and that their susceptibility depends on the severity of hypercholesterolemia. Wild-type (WT) mice and ApoE(-/-) mice fed a low-cholesterol (LC) or high-cholesterol (HC) diet were infected with approximately 50 CFU Mycobacterium tuberculosis Erdman by aerosol. ApoE(-/-) LC mice were modestly more susceptible to tuberculosis than WT LC mice. In contrast, ApoE(-/-) HC mice were extremely susceptible, as evidenced by 100% mortality after 4 weeks with tuberculosis. The lung pathology of ApoE(-/-) HC mice was remarkable for giant abscess-like lesions, massive infiltration by granulocytes, elevated inflammatory cytokine production, and a mean bacterial load approximately 2 log units higher than that of WT HC mice. Compared to WT HC mice, the gamma interferon response of splenocytes restimulated ex vivo with M. tuberculosis culture filtrate protein was delayed in ApoE(-/-) HC mice, and they failed to control M. tuberculosis growth in the lung. OT-II cells adoptively transferred into uninfected ApoE(-/-) HC mice had a weak proliferative response to their antigen, indicating impaired priming of the adaptive immune response. Our studies show that ApoE(-/-) deficiency is associated with delayed expression of adaptive immunity to tuberculosis caused by defective priming of the adaptive immune response and that elevated serum cholesterol is responsible for this effect.
Figures

Comment in
-
Is hypercholesterolemia a friend or a foe of tuberculosis?Infect Immun. 2009 Aug;77(8):3514; author reply 3514-5. doi: 10.1128/IAI.00469-09. Infect Immun. 2009. PMID: 19617455 Free PMC article. No abstract available.
References
-
- Alieke, G. V., N. de Bont, M. G. Netea, P. N. Demacker, J. W. van der Meer, A. F. Stalenhoef, and B. J. Kullberg. 2004. Apolipoprotein-E-deficient mice exhibit an increased susceptibility to disseminated candidiasis. Med. Mycol. 42341-348. - PubMed
-
- Angeli, V., J. Llodra, J. X. Rong, K. Satoh, S. Ishii, T. Shimizu, E. A. Fisher, and G. J. Randolph. 2004. Dyslipidemia associated with atherosclerotic disease systemically alters dendritic cell mobilization. Immunity 21561-574. - PubMed
-
- Barnden, M. J., J. Allison, W. R. Heath, and F. R. Carbone. 1998. Defective TCR expression in transgenic mice constructed using cDNA-based alpha- and beta-chain genes under the control of heterologous regulatory elements. Immunol. Cell Biol. 7634-40. - PubMed
-
- Bjorkbacka, H., V. V. Kunjathoor, K. J. Moore, S. Koehn, C. M. Ordija, M. A. Lee, T. Means, K. Halmen, A. D. Luster, D. T. Golenbock, and M. W. Freeman. 2004. Reduced atherosclerosis in MyD88-null mice links elevated serum cholesterol levels to activation of innate immunity signaling pathways. Nat. Med. 10416-421. - PubMed
-
- de Bont, N., M. G. Netea, P. N. Demacker, B. J. Kullberg, J. W. van der Meer, and A. F. Stalenhoef. 2000. Apolipoprotein E-deficient mice have an impaired immune response to Klebsiella pneumoniae. Eur. J. Clin. Investig. 30818-822. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
