

p53 stabilization in response to DNA damage requires Akt/PKB and DNA-PK
- PMID: 18505846
- PMCID: PMC2409394
- DOI: 10.1073/pnas.0703423105
p53 stabilization in response to DNA damage requires Akt/PKB and DNA-PK
Abstract
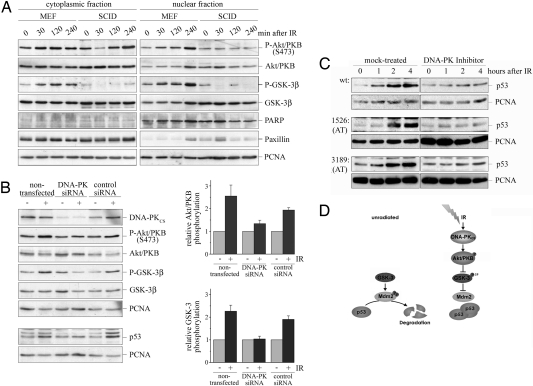
The p53 protein is one of the major tumor suppressor proteins. In response to DNA damage, p53 is prevented from degradation and accumulates to high levels. Ionizing radiation leads to hypophosphorylation of the p53 ubiquitin ligase Mdm2 at sites where phosphorylation is critical for p53 degradation and to the phosphorylation and activation of Akt/PKB, a kinase that phosphorylates and inhibits GSK-3. GSK-3, which normally phosphorylates Mdm2, is inactivated in response to ionizing radiation. We show that p53 accumulates in lymphoblasts from patients with the hereditary disorder ataxia telangiectasia in response to ionizing radiation despite the absence of a functional ATM kinase. Also, knockdown of ATR did not prevent p53 accumulation in response to ionizing radiation. Instead, p53 stabilization in response to ionizing radiation depended on the inactivation of GSK-3 and the presence of Akt/PKB. Akt/PKB is a target of DNA-PK, a kinase that is activated after ionizing radiation. Correspondingly, down-regulation of DNA-PK prevented phosphorylation of Akt/PKB and GSK-3 after ionizing radiation and strongly reduced the accumulation of p53. We therefore propose a signaling cascade for the regulation of p53 in response to ionizing radiation that involves activation of DNA-PK and Akt/PKB and inactivation of GSK-3 and Mdm2.
Conflict of interest statement
The authors declare no conflict of interest.
Figures




References
-
- Gasser S, Raulet D. The DNA damage response, immunity and cancer. Semin Cancer Biol. 2006;5:344–347. - PubMed
-
- Donehower LA, et al. Deficiency of p53 accelerates mammary tumorigenesis in Wnt-1 transgenic mice and promotes chromosomal instability. Genes Dev. 1995;9:882–895. - PubMed
-
- Levine AJ, Hu W, Feng Z. The p53 pathway: What questions remain to be explored? Cell Death Differ. 2006;13:1027–1036. - PubMed
-
- Lavin MF, Gueven N. The complexity of p53 stabilization and activation. Cell Death Differ. 2006;6:941–950. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Research Materials
Miscellaneous
