

Induction of plasma (TRAIL), TNFR-2, Fas ligand, and plasma microparticles after human immunodeficiency virus type 1 (HIV-1) transmission: implications for HIV-1 vaccine design
- PMID: 18508902
- PMCID: PMC2493338
- DOI: 10.1128/JVI.00605-08
Induction of plasma (TRAIL), TNFR-2, Fas ligand, and plasma microparticles after human immunodeficiency virus type 1 (HIV-1) transmission: implications for HIV-1 vaccine design
Abstract
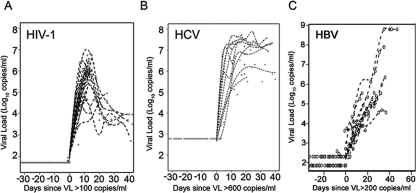
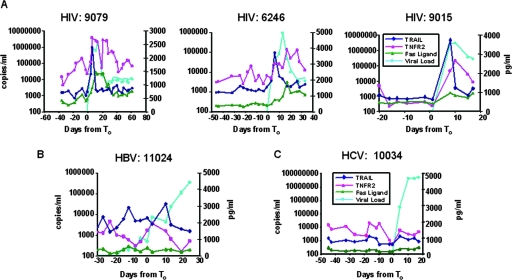
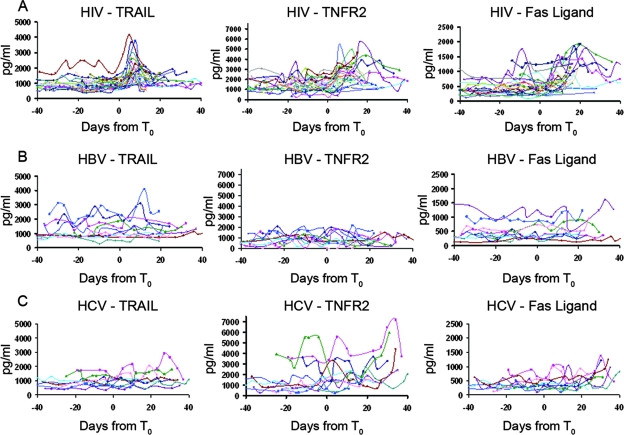
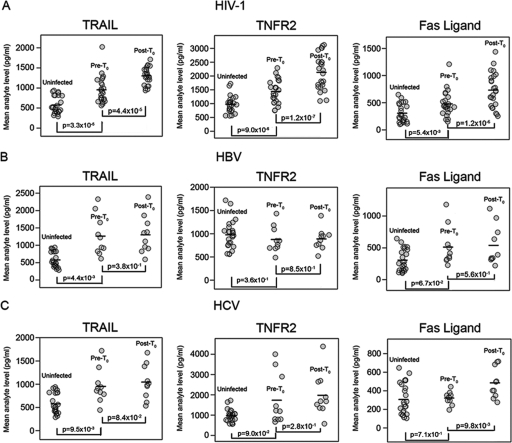
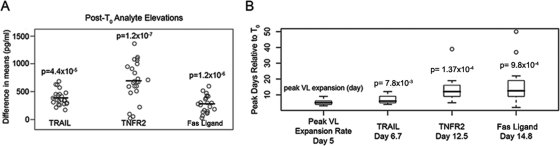
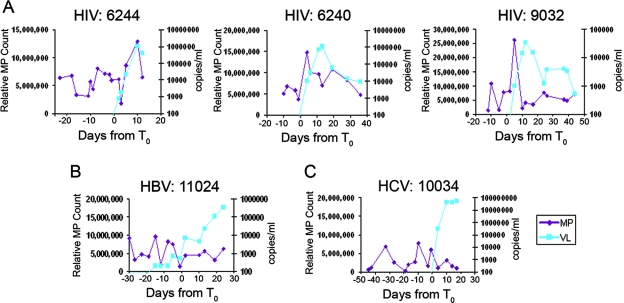
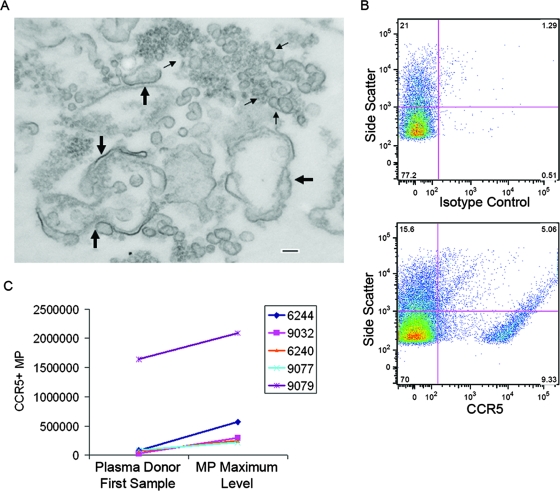
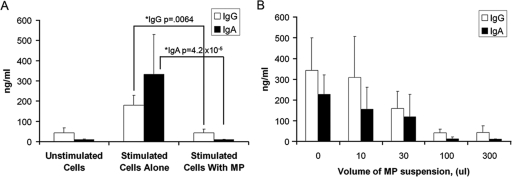
The death of CD4(+) CCR5(+) T cells is a hallmark of human immunodeficiency virus (HIV) infection. We studied the plasma levels of cell death mediators and products--tumor necrosis factor (TNF)-related apoptosis-inducing ligand (TRAIL), Fas ligand, TNF receptor type 2 (TNFR-2), and plasma microparticles--during the earliest stages of infection following HIV type 1 (HIV-1) transmission in plasma samples from U.S. plasma donors. Significant plasma TRAIL level elevations occurred a mean of 7.2 days before the peak of plasma viral load (VL), while TNFR-2, Fas ligand, and microparticle level elevations occurred concurrently with maximum VL. Microparticles had been previously shown to mediate immunosuppressive effects on T cells and macrophages. We found that T-cell apoptotic microparticles also potently suppressed in vitro immunoglobulin G (IgG) and IgA antibody production by memory B cells. Thus, release of TRAIL during the onset of plasma viremia (i.e., the eclipse phase) in HIV-1 transmission may initiate or amplify early HIV-1-induced cell death. The window of opportunity for a HIV-1 vaccine is from the time of HIV-1 transmission until establishment of the latently infected CD4(+) T cells. Release of products of cell death and subsequent immunosuppression following HIV-1 transmission could potentially narrow the window of opportunity during which a vaccine is able to extinguish HIV-1 infection and could place severe constraints on the amount of time available for the immune system to respond to the transmitted virus.
Figures
References
-
- Admyre, C., S. M. Johansson, K. R. Qazi, J. J. Filén, R. Lahesmaa, M. Norman, E. P. Neve, A. Scheynius, and S. Gabrielsson. 2007. Exosomes with immune modulatory features are present in human breast milk. J. Immunol. 1791969-1978. - PubMed
-
- Aukrust, P., N. B. Liabakk, F. Muller, E. Lien, T. Espevik, and S. S. Froland. 1994. Serum levels of tumor necrosis factor-alpha (TNF alpha) and soluble TNF receptors in human immunodeficiency virus type 1 infection—correlations to clinical, immunologic, and virologic parameters. J. Infect. Dis. 169420-424. - PubMed
-
- Badley, A. D., A. A. Pilon, A. Landay, and D. H. Lynch. 2000. Mechanisms of HIV-associated lymphocyte apoptosis. Blood 962951-2964. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Research Materials
Miscellaneous
