

Xenopus TACC3/maskin is not required for microtubule stability but is required for anchoring microtubules at the centrosome
- PMID: 18508920
- PMCID: PMC2488304
- DOI: 10.1091/mbc.e07-11-1204
Xenopus TACC3/maskin is not required for microtubule stability but is required for anchoring microtubules at the centrosome
Abstract
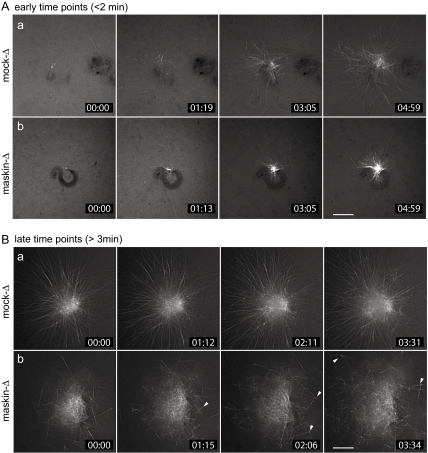
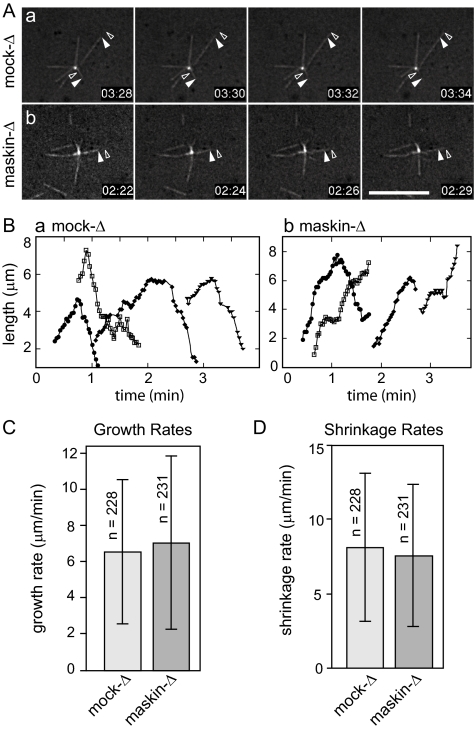
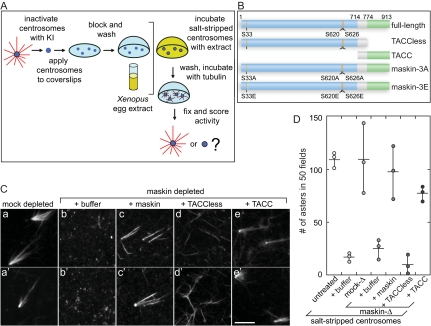
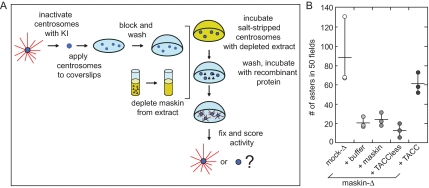
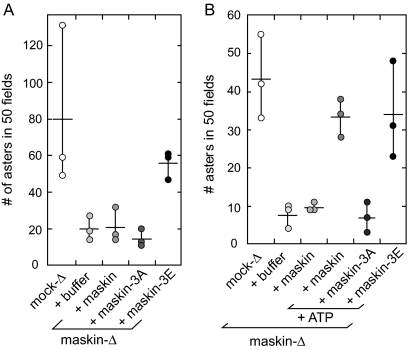
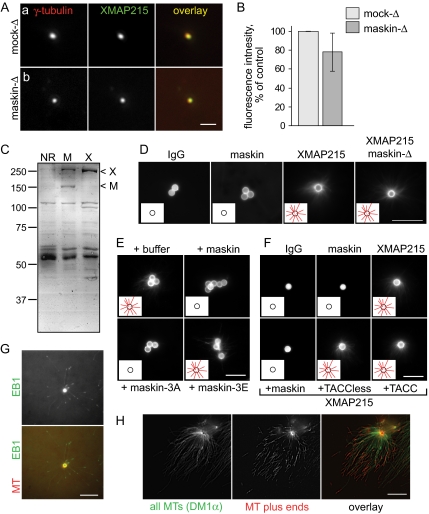
Members of the transforming acidic coiled coil (TACC) protein family are emerging as important mitotic spindle assembly proteins in a variety of organisms. The molecular details of how TACC proteins function are unknown, but TACC proteins have been proposed to recruit microtubule-stabilizing proteins of the tumor overexpressed gene (TOG) family to the centrosome and to facilitate their loading onto newly emerging microtubules. Using Xenopus egg extracts and in vitro assays, we show that the Xenopus TACC protein maskin is required for centrosome function beyond recruiting the Xenopus TOG protein XMAP215. The conserved C-terminal TACC domain of maskin is both necessary and sufficient to restore centrosome function in maskin-depleted extracts, and we provide evidence that the N terminus of maskin inhibits the function of the TACC domain. Time-lapse video microscopy reveals that microtubule dynamics in Xenopus egg extracts are unaffected by maskin depletion. Our results provide direct experimental evidence of a role for maskin in centrosome function and suggest that maskin is required for microtubule anchoring at the centrosome.
Figures
References
-
- Albee A. J., Tao W., Wiese C. Phosphorylation of maskin by Aurora-A is regulated by RanGTP and importin β. J. Biol. Chem. 2006;281:38293–38301. - PubMed
-
- Andersen J. S., Wilkinson C. J., Mayor T., Mortensen P., Nigg E. A., Mann M. Proteomic characterization of the human centrosome by protein correlation profiling. Nature. 2003;426:570–574. - PubMed
-
- Azimzadeh J., Bornens M. Structure and duplication of the centrosome. J. Cell Sci. 2007;120:2139–2142. - PubMed
-
- Badano J. L., Teslovich T. M., Katsanis N. The centrosome in human genetic disease. Nat. Rev. Genet. 2005;6:194–205. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
