

Social interaction-mediated lifespan extension of Drosophila Cu/Zn superoxide dismutase mutants
- PMID: 18508973
- PMCID: PMC2396722
- DOI: 10.1073/pnas.0711127105
Social interaction-mediated lifespan extension of Drosophila Cu/Zn superoxide dismutase mutants
Abstract
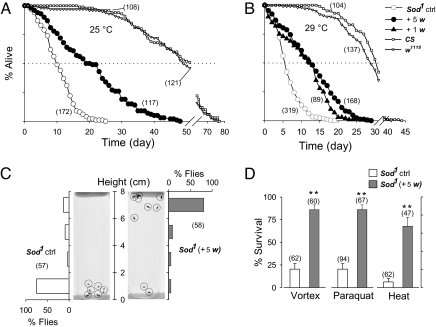
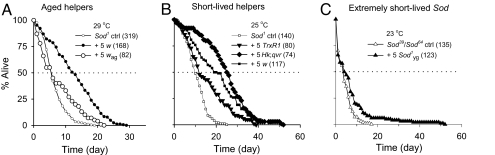
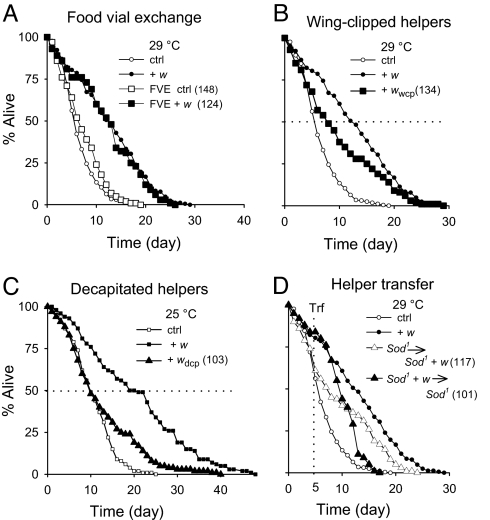
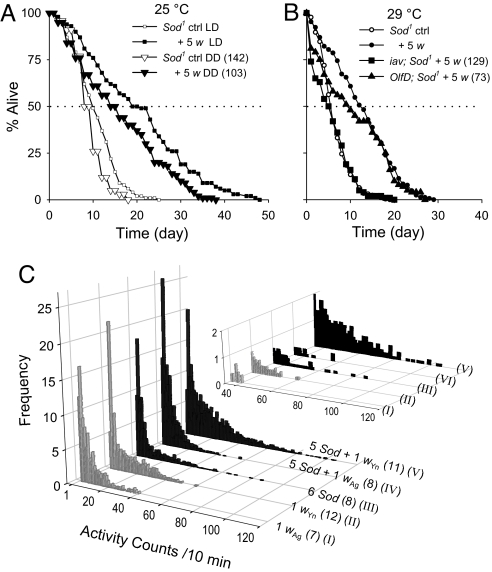
Beneficial effects of social interaction on aging have been studied in humans and other species. We found that short-lived Drosophila mutants of the antioxidant enzyme Cu/Zn superoxide dismutase displayed a robust lifespan extension, with improved stress resistance and motor ability, upon cohousing with active flies of longer lifespan or younger age. Genetic, surgical, and environmental manipulations revealed motor and sensory components in behavioral interactions required for the lifespan extension induced by cohousing. Our results provide a definitive case of beneficial social interaction on lifespan and a useful entry point for analyzing the underlying molecular networks and physiological mechanisms.
Conflict of interest statement
The authors declare no conflict of interest.
Figures
References
-
- Nee LE. Effects of psychosocial interactions at a cellular level. Soc Work. 1995;40:259–262. - PubMed
-
- Mountz JD, Zant GV, Allison DB, Zhang HG, Hsu HC. Beneficial influences of systemic cooperation and sociological behavior on longevity. Mech Ageing Dev. 2002;123:963–973. - PubMed
-
- Sapolsky RM. The influence of social hierarchy on primate health. Science. 2005;308:648–652. - PubMed
-
- Hennessy MB. Social influences on endocrine activity in guinea pigs, with comparisons to findings in nonhuman primates. Neurosci Biobehav Rev. 1999;23:687–698. - PubMed
-
- Menich SR, Baron A. Social housing of rats: Life-span effects on reaction time, exploration, weight, and longevity. Exp Aging Res. 1984;10:95–100. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
