

Arousal facilitates collision avoidance mediated by a looming sensitive visual neuron in a flying locust
- PMID: 18509080
- PMCID: PMC2525709
- DOI: 10.1152/jn.01055.2007
Arousal facilitates collision avoidance mediated by a looming sensitive visual neuron in a flying locust
Abstract
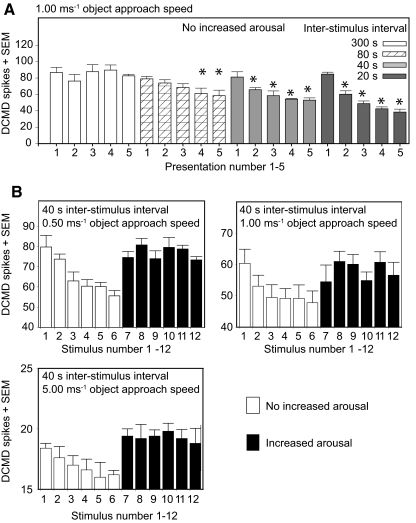
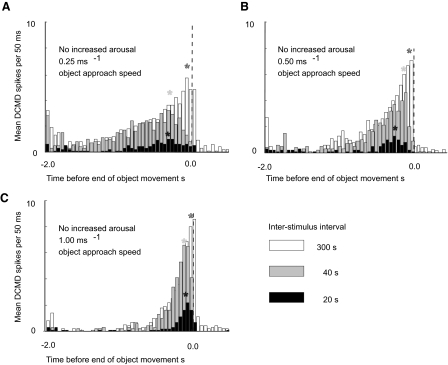
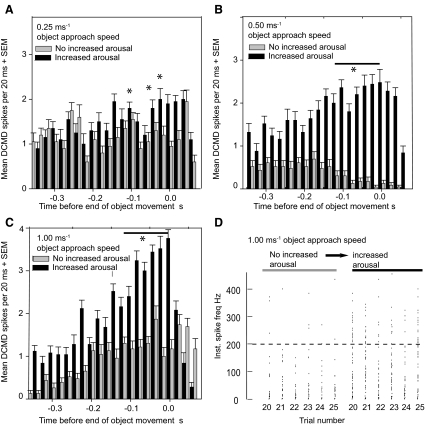
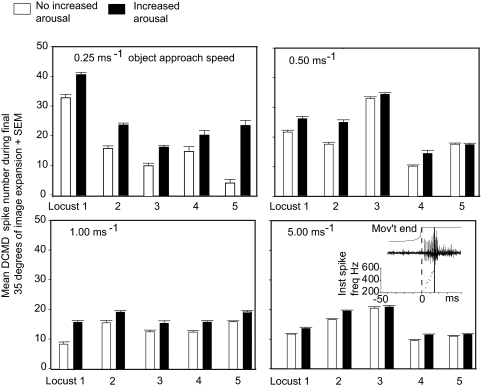
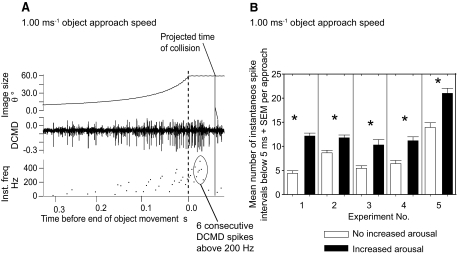
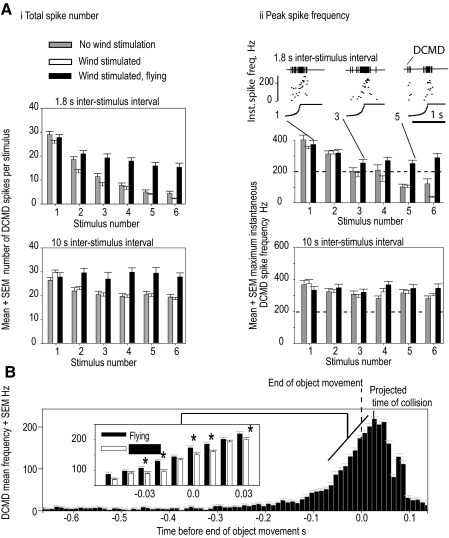
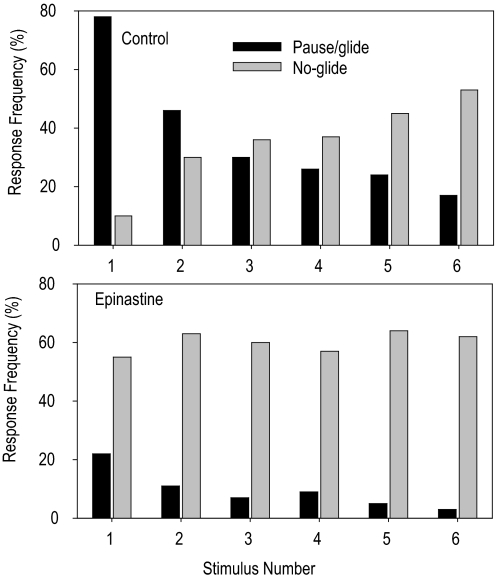
Locusts have two large collision-detecting neurons, the descending contralateral movement detectors (DCMDs) that signal object approach and trigger evasive glides during flight. We sought to investigate whether vision for action, when the locust is in an aroused state rather than a passive viewer, significantly alters visual processing in this collision-detecting pathway. To do this we used two different approaches to determine how the arousal state of a locust affects the prolonged periods of high-frequency spikes typical of the DCMD response to approaching objects that trigger evasive glides. First, we manipulated arousal state in the locust by applying a brief mechanical stimulation to the hind leg; this type of change of state occurs when gregarious locusts accumulate in high-density swarms. Second, we examined DCMD responses during flight because flight produces a heightened physiological state of arousal in locusts. When arousal was induced by either method we found that the DCMD response recovered from a previously habituated state; that it followed object motion throughout approach; and--most important--that it was significantly more likely to generate the maintained spike frequencies capable of evoking gliding dives even with extremely short intervals (1.8 s) between approaches. Overall, tethered flying locusts responded to 41% of simulated approaching objects (sets of 6 with 1.8 s ISI). When we injected epinastine, the neuronal octopamine receptor antagonist, into the hemolymph responsiveness declined to 12%, suggesting that octopamine plays a significant role in maintaining responsiveness of the DCMD and the locust to visual stimuli during flight.
Figures
References
-
- Araki M, Nagayama T. Direct chemically mediated synaptic transmission from mechanosensory afferents contributes to habituation of crayfish lateral giant escape reaction. J Comp Physiol A Sens Neural Behav Physiol 189: 731–739, 2003. - PubMed
-
- Araki M, Nagayama T, Sprayberry J. Cyclic AMP mediates serotonin-induced synaptic enhancement of lateral giant interneuron of the crayfish. J Neurophysiol 94: 2644–2652, 2005. - PubMed
-
- Aston-Jones G, Chiang C, Alexinsky T. Discharge of noradrenergic locus coeruleus neurons in behaving rats and monkeys suggests a role in vigilance. Prog Brain Res 88: 501–520, 1992. - PubMed
-
- Bacon JP, Thompson KSJ, Stern M. Identified octopaminergic neurons provide an arousal mechanism in the locust brain. J Neurophysiol 74: 2739–2743, 1995. - PubMed
-
- Barron AB, Maleszka J,. Van der Meer RK, Robinson GE, Maleszka R. Comparing injection, feeding and topical application methods for treatment of honey bees with octopamine. J Insect Physiol 53: 187–194, 2007. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources