

The interaction network of the chaperonin CCT
- PMID: 18511909
- PMCID: PMC2486426
- DOI: 10.1038/emboj.2008.108
The interaction network of the chaperonin CCT
Abstract
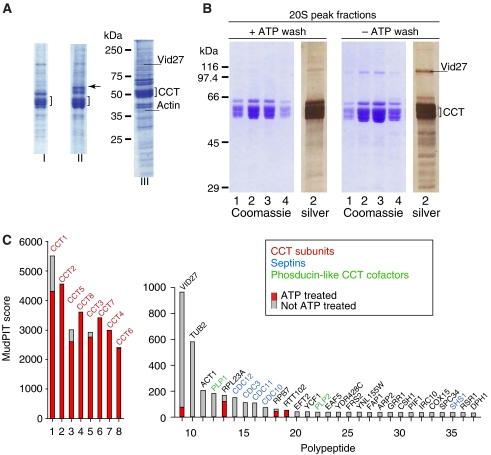
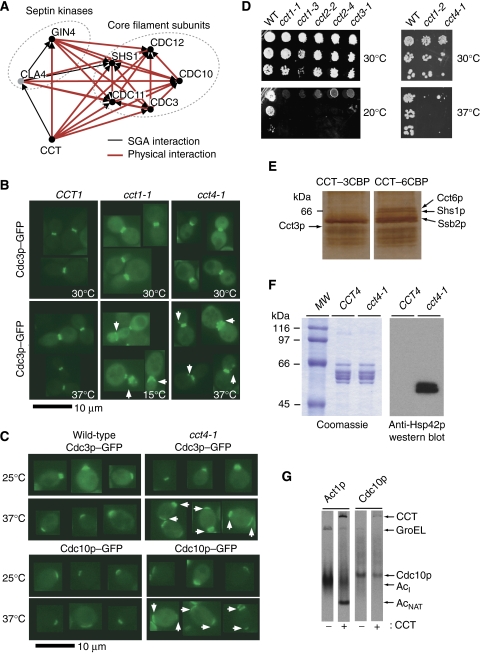
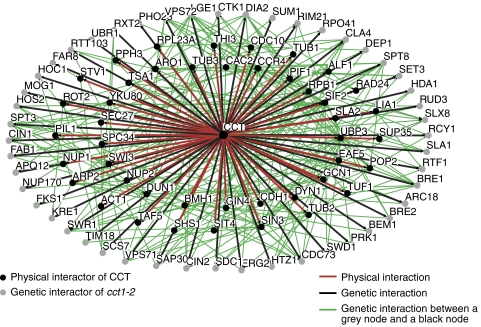
The eukaryotic cytosolic chaperonin containing TCP-1 (CCT) has an important function in maintaining cellular homoeostasis by assisting the folding of many proteins, including the cytoskeletal components actin and tubulin. Yet the nature of the proteins and cellular pathways dependent on CCT function has not been established globally. Here, we use proteomic and genomic approaches to define CCT interaction networks involving 136 proteins/genes that include links to the nuclear pore complex, chromatin remodelling, and protein degradation. Our study also identifies a third eukaryotic cytoskeletal system connected with CCT: the septin ring complex, which is essential for cytokinesis. CCT interactions with septins are ATP dependent, and disrupting the function of the chaperonin in yeast leads to loss of CCT-septin interaction and aberrant septin ring assembly. Our results therefore provide a rich framework for understanding the function of CCT in several essential cellular processes, including epigenetics and cell division.
Figures

References
-
- Adams A, Gottschling DE, Kaiser CA, Stearns T (1997) Methods in Yeast Genetics: a Cold Spring Harbor Laboratory Course Manual. Cold Spring Harbour, New York, USA: Cold Spring Harbour Laboratory Press
-
- Aloy P, Böttcher B, Ceulemans H, Leutwein C, Mellwig C, Fischer S, Gavin AC, Bork P, Superti-Furga G, Serrano L, Russell RB (2004) Structure-based assembly of protein complexes in yeast. Science 303: 2026–2029 - PubMed
-
- Amberg DC, Burke DJ, Strathern JN (2005) Methods in Yeast Genetics. Cold Spring Harbour, New York, USA: Cold Spring Harbour Laboratory Press
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
