

The Campylobacter jejuni thiol peroxidases Tpx and Bcp both contribute to aerotolerance and peroxide-mediated stress resistance but have distinct substrate specificities
- PMID: 18515414
- PMCID: PMC2493253
- DOI: 10.1128/JB.00100-08
The Campylobacter jejuni thiol peroxidases Tpx and Bcp both contribute to aerotolerance and peroxide-mediated stress resistance but have distinct substrate specificities
Abstract
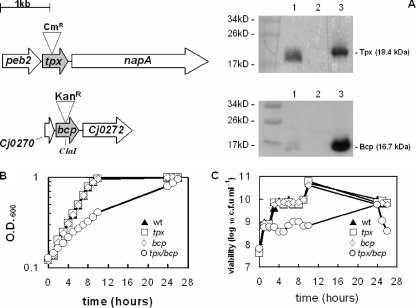
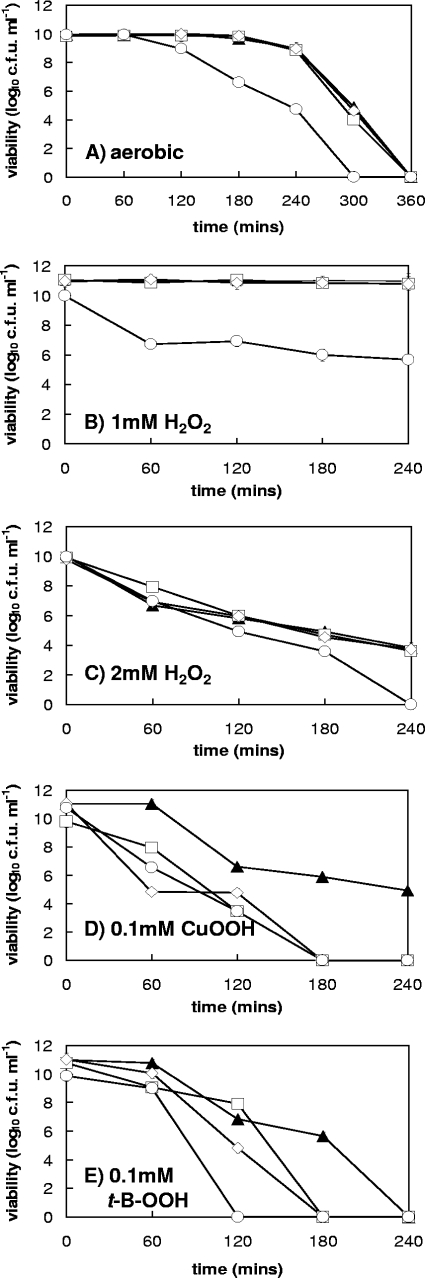
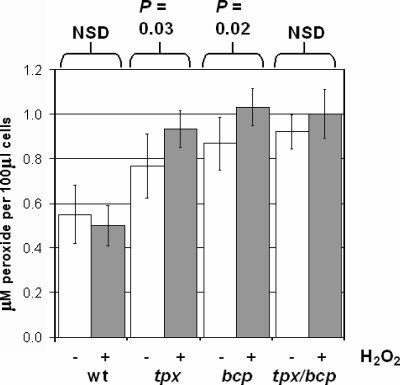
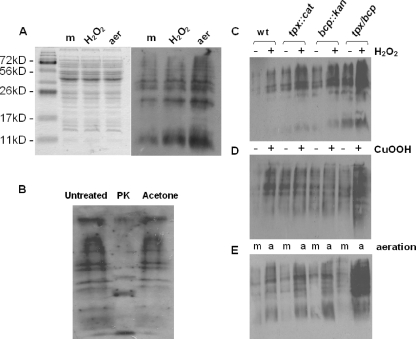
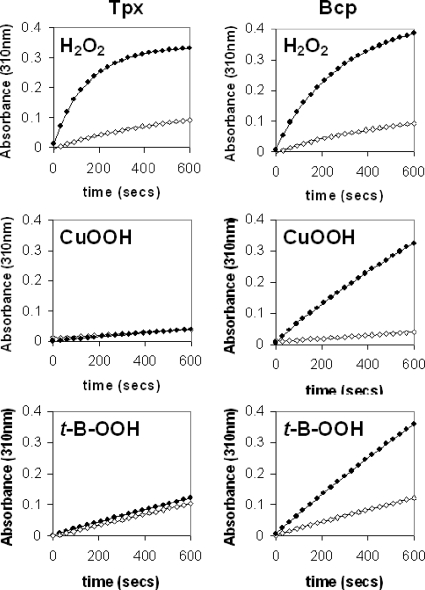
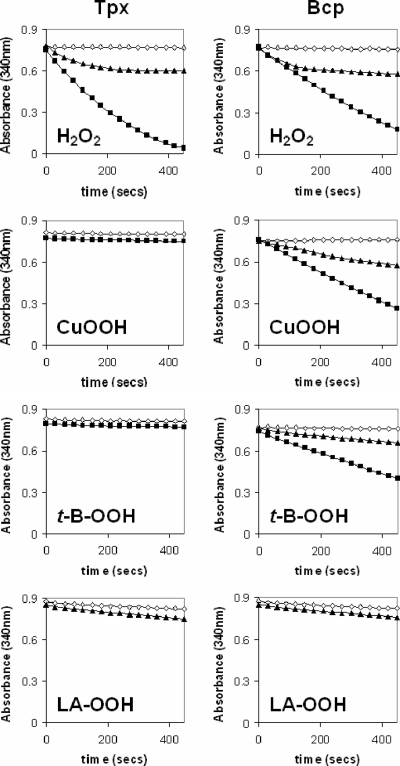
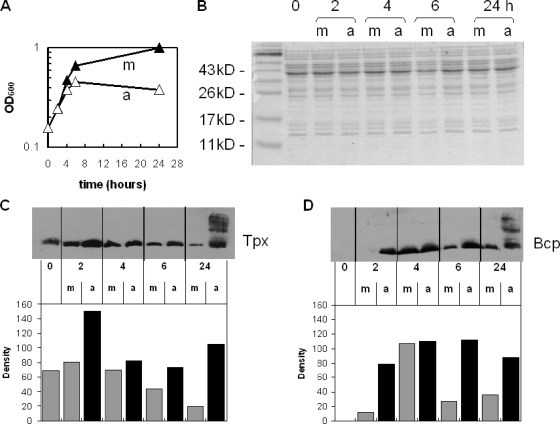
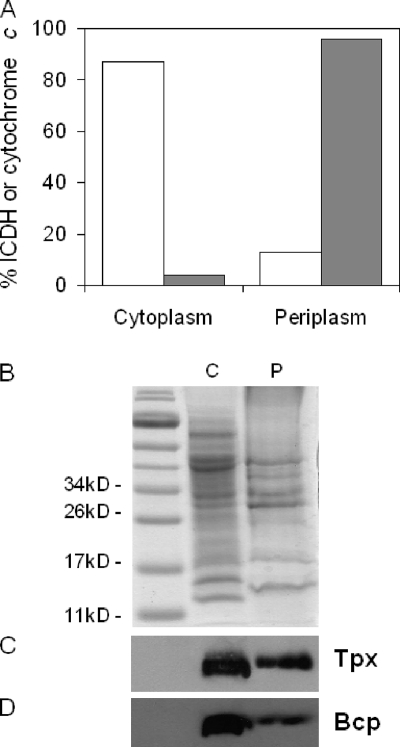
The microaerophilic food-borne pathogen Campylobacter jejuni experiences variable oxygen concentrations during its life cycle, especially during transitions between the external environment and the avian or mammalian gut. Single knockout mutations in either one of two related thiol peroxidase genes, tpx and bcp, resulted in normal microaerobic growth (10% [vol/vol] oxygen) but poorer growth than that of the wild type under high-aeration conditions (21% [vol/vol] oxygen). However, a tpx/bcp double mutant had a severe microaerobic growth defect and did not grow at high aeration in shake flasks. Although the single mutant strains were no more sensitive than the wild-type strains in disc diffusion assays with hydrogen peroxide, organic peroxides, superoxide, or nitrosative stress agents, in all cases the double mutant was hypersensitive. Quantitative cell viability and cellular lipid peroxidation assays indicated some increased sensitivity of the single tpx and bcp mutants to peroxide stress. Protein carbonylation studies revealed that the tpx/bcp double mutant had a higher degree of oxygen- and peroxide-induced oxidative protein damage than did either of the single mutants. An analysis of the peroxidase activity of the purified recombinant enzymes showed that, surprisingly, Tpx reduced only hydrogen peroxide as substrate, whereas Bcp also reduced organic peroxides. Immunoblotting of wild-type cell extracts with Tpx- or Bcp-specific antibodies showed increased abundance of both proteins under high aeration compared to that under microaerobic growth conditions. Taken together, the results suggest that Tpx and Bcp are partially redundant antioxidant enzymes that play an important role in protection of C. jejuni against oxygen-induced oxidative stress.
Figures
References
-
- Atack, J. M., and D. J. Kelly. 2006. Structure, mechanism and physiological roles of bacterial cytochrome c peroxidases. Adv. Microb. Physiol. 5273-106. - PubMed
-
- Bryk, R., P. Griffin, and C. Nathan. 2000. Peroxynitrite reductase activity of bacterial peroxiredoxins. Nature 407211-215. - PubMed
-
- Cha, M., H. Kim, and I. Kim. 1995. Thioredoxin-linked “thiol peroxidase” from periplasmic space of Escherichia coli. J. Biol. Chem. 27028635-28641. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
