

Leptin controls adipose tissue lipogenesis via central, STAT3-independent mechanisms
- PMID: 18516053
- PMCID: PMC2671848
- DOI: 10.1038/nm1775
Leptin controls adipose tissue lipogenesis via central, STAT3-independent mechanisms
Abstract
Leptin (encoded by Lep) controls body weight by regulating food intake and fuel partitioning. Obesity is characterized by leptin resistance and increased endocannabinoid tone. Here we show that leptin infused into the mediobasal hypothalamus (MBH) of rats inhibits white adipose tissue (WAT) lipogenesis, which occurs independently of signal transducer and activator of transcription-3 (STAT3) signaling. Correspondingly, transgenic inactivation of STAT3 signaling by mutation of the leptin receptor (s/s mice) leads to reduced adipose mass compared to db/db mice (complete abrogation of leptin receptor signaling). Conversely, the ability of hypothalamic leptin to suppress WAT lipogenesis in rats is lost when hypothalamic phosphoinositide 3-kinase signaling is prevented or when sympathetic denervation of adipose tissue is performed. MBH leptin suppresses the endocannabinoid anandamide in WAT, and, when this suppression of endocannabinoid tone is prevented by systemic CB1 receptor activation, MBH leptin fails to suppress WAT lipogenesis. These data suggest that the increased endocannabinoid tone observed in obesity is linked to a failure of central leptin signaling to restrain peripheral endocannabinoids.
Figures
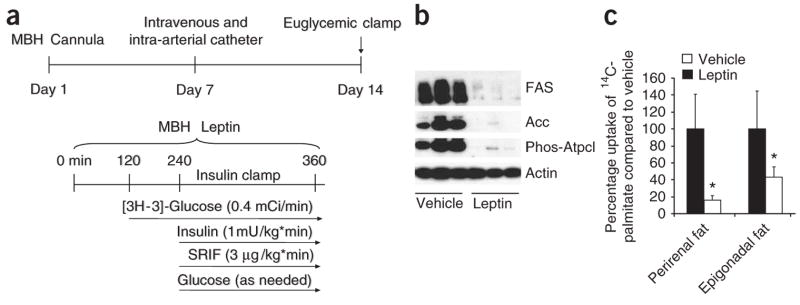
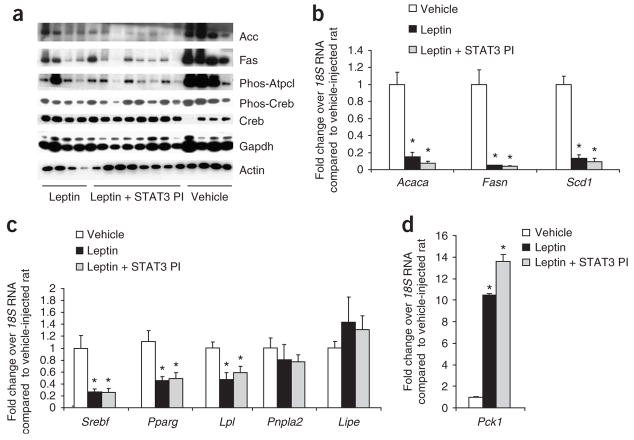
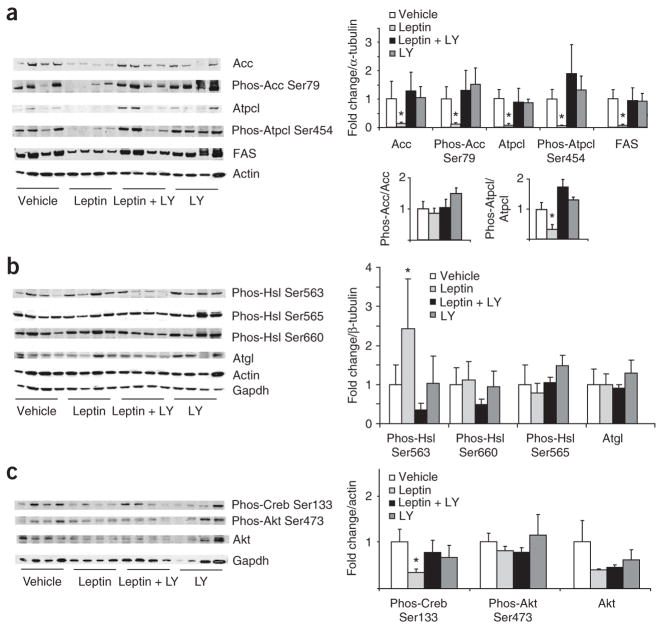
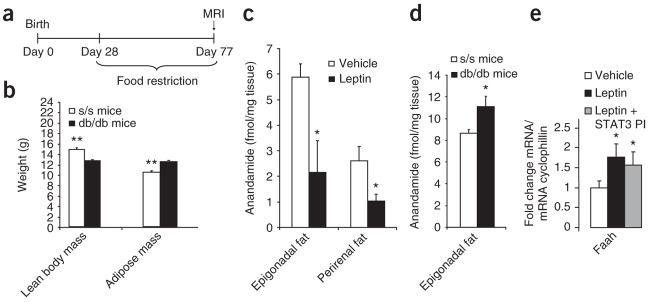
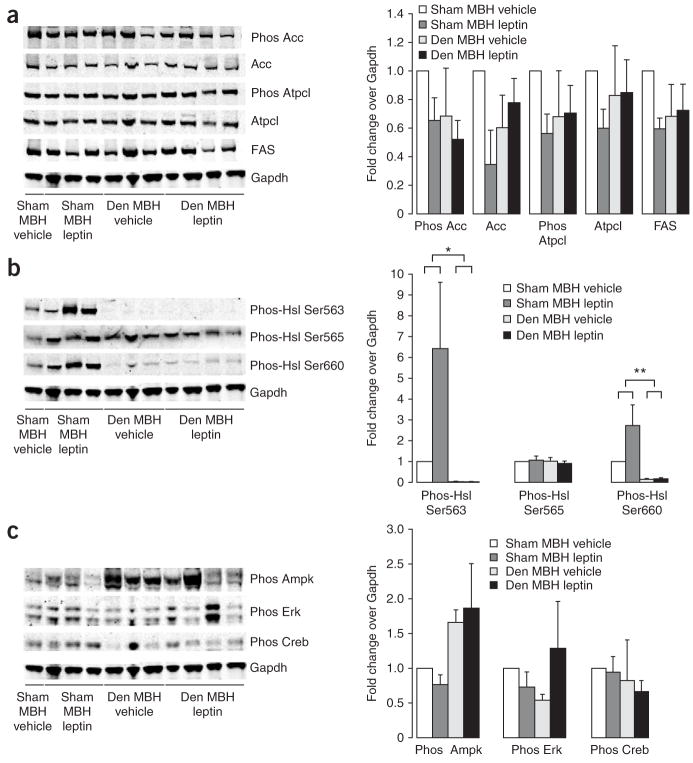
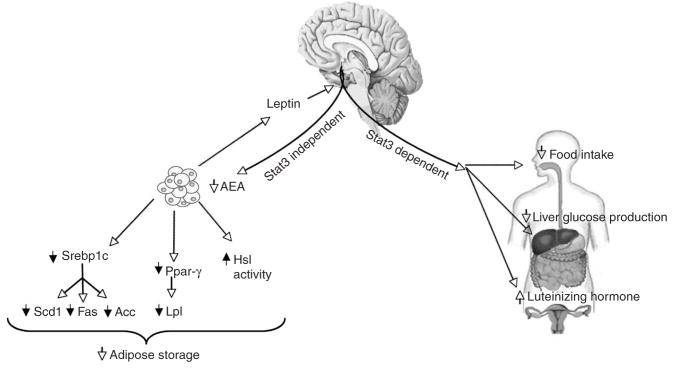
Comment in
-
The long reach of leptin.Nat Med. 2008 Jun;14(6):604-6. doi: 10.1038/nm0608-604. Nat Med. 2008. PMID: 18535571 No abstract available.
References
-
- Matias I, et al. Regulation, function, and dysregulation of endocannabinoids in models of adipose and β-pancreatic cells and in obesity and hyperglycemia. J Clin Endocrinol Metab. 2006;91:3171–3180. - PubMed
-
- Woods SC. The endocannabinoid system: mechanisms behind metabolic homeostasis and imbalance. Am J Med. 2007;120:S9–S17. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
